

Cała niniejsza praca jest reakcją na pojawienie się i uporczywe rozpowszechnianie w środowisku narodowym ideologii obywatelskiego nacjonalizmu. Odpowiedź autora tego artykułu na to zjawisko ideologiczne rozpoczęła się od przedstawienia w pierwszej części tego eseju źródeł ideologicznych obywatelskiego nacjonalizmu, tkwiących w opozycyjnych wobec indoeuropejskiej tradycji, ideach oświecenia. Następnie, pokrótce została przedstawiona na przestrzeni czasu próba naukowego umocowania oświeceniowych idei, która po przejściowym okresie sporego powodzenia zakończyła się ostateczną klęską. Niepowodzenie naukowego uzasadnienia aksjomatów oświeceniowej rewolty wobec tradycji było spowodowane powstaniem nowych gałęzi nauki, bazujących na matematycznej teorii gier, genetyce oraz ewolucyjnych naukach społecznych i psychologicznych z jednej strony, oraz na udowodnieniu fałszywości rzekomych „ustaleń naukowych” poczynionych przez zaangażowanych ideologicznie lewicowców, którzy udawali naukowych badaczy. Kolejnym krokiem autora tego eseju, służącym ukazaniu absurdalności postawy obywatelskiej wśród polskich narodowców, było prześledzenie powstania i rozwoju nowych dyscyplin naukowych, bazujących na teorii ewolucji K. Darwina oraz na naukach ścisłych. Przełomowe znaczenie tego procesu polega bowiem na tym, że traktowane dotychczas intuicyjnie kategorie narodu, patriotyzmu, solidaryzmu narodowego, uzyskały w ramach tych dyscyplin wiedzy naukową konceptualizację, oraz stały się kwantyfikowalne.
Etniczny nacjonalizm zyskał więc trzy solidne fundamenty naukowe, które miażdżą ogromem danych empirycznych stanowisko „obywatelskie” jako zwykłą umysłową aberrację. Dwa z tych fundamentów czyli matematyczna teoria gier oraz teoria podobieństwa genetycznego (GST) zostały omówione (wraz z krytyką) zamykając pierwszą część.
Tym razem autor zajmie się koncepcją interesu genetycznego, stworzoną przez F. Saltera w jego opus magnum pt. „On genetic interests. Family, ethnicity and humanity in an age of mass migration”.
Jak to zostało ustalone w poprzednich tekstach, podstawowy cel życia zawiera się w imperatywie „przetrwaj aż do rozmnożenia”. Rozmnażanie jest niczym innym jak replikowaniem własnego projektu genetycznego w kolejnych generacjach w czasie. Jednak bezpośrednie klonowanie dotyczy jedynie bardzo prostych form życia. Człowiek, jak większość gatunków stojących wysoko w hierarchii ewolucyjnej, rozmnaża się płciowo, co zapewnia zróżnicowanie genetyczne potrzebne dla ciągłej ewolucji. Jak wiemy z teorii Hamiltona o dostosowaniu łącznym, sukces reprodukcyjny danego osobnika w obrębie gatunku zależy częściowo od sukcesu reprodukcyjnego innych osób, które dzielą niektóre z jego charakterystycznych genów. Oznacza to, że dla selekcji naturalnej liczy się nie tylko bezpośrednie rodzicielstwo ale także rodzina w sensie rozszerzonym.
Zatem interes genetyczny danej osoby w określonej grupie (ród, plemię, naród), to liczba kopii charakterystycznych genów tej osoby przenoszonych w tej grupie z pokolenia na pokolenie poprzez rozmnażanie się jej członków. Interesy genetyczne jednostki w grupie mają dwa wymiary: po pierwsze średnie stężenie genów danej jednostki w grupie na osobę, oraz po drugie, wielkość grupy.
Kiedy zaczynamy intuicyjnie myśleć o jakiś charakterystycznych cechach danych ludzi, od razu nasuwa nam się kategoria rodziny, w której w zależności od stopnia pokrewieństwa możemy określać dziedziczenie konkretnych cech. Myślenie to pokrywa się zasadniczo z koncepcją Hamiltona, która opiera się właśnie na stopniu pokrewieństwa w którym odsetek wspólnych genów oznaczany w równaniach jako r, (od słowa relatedness - pokrewieństwo) zmniejsza się o 50% co każdy stopień pokoleniowy. Jednostka w tym modelu dzieli połowę swoich genów z dziećmi, ćwierć genów z wnukami, ósmą część z kuzynami itp. Prostym przykładem takiej kwantyfikacji jest przypadek pokrewieństwa między kuzynami.
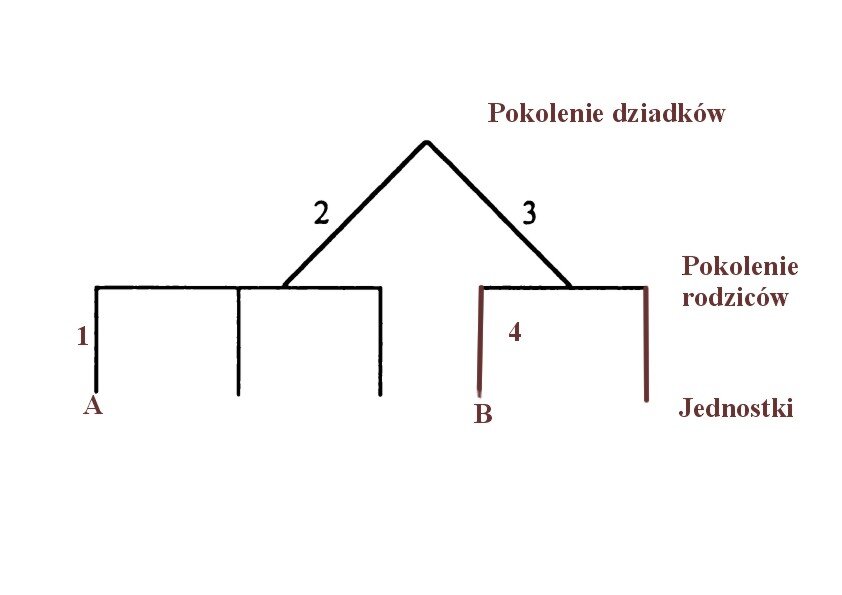
Na rysunku widzimy stopnie pokrewieństwa pomiędzy dwoma kuzynami. Metoda polega na policzeniu kroków pokoleniowych od jednej osoby do wspólnego przodka a następnie, od wspólnego przodka do drugiej osoby. Jak pamiętamy pokrewieństwo zmniejsza się o połowę co każdy stopień pokoleniowy. W tym przypadku
r= 0,54 = 0,0625
Jako, że istnieją cztery kroki pokoleniowe, stopień pokrewieństwa jest równy połowie podniesionej do czwartej potęgi czyli jedna szesnasta. Jednak ponieważ kuzyni w pierwszym pokoleniu dzielą dwóch dziadków wynik należy pomnożyć przez dwa co daje w sumie pokrewieństwo jednej ósmej (0, 125).
W nawiązaniu do definicji interesu genetycznego, przykładowa rodzina posiadająca trójkę dzieci będzie posiadać w ich pokoleniu 1,5 kopi każdego z rodziców, w kolejnym pokoleniu siostrzenice i siostrzeńcy dadzą kolejne 1,5 kopi genów (6 x 0,25) a kuzyni w pierwszym pokoleniu kolejne 1,5 kopi (6 x 0,25). Razem, te trzy zestawy krewnych dadzą 4,5 kopi genów każdego z rodziców. W takim samym zestawie jeżeli w każdym pokoleniu będzie dwoje dzieci to taki zestaw ma 2 kopie genów każdego z rodziców, jeżeli zaś dzieci będzie czworo to kopii rodziców w pokoleniu kuzynów będzie 6. Widać więc wyraźnie, że przy stałym dziedziczeniu na poziomie 0.5 drugim czynnikiem zwiększającym interes genetyczny jednostki jest ilość krewnych, która zwiększa liczebnie grupę. Tak więc im dalej poza rodzinny horyzont współczesnej rodziny, przy dużej liczbie ludności, wspólni przodkowie będą mieli więcej kopii swoich charakterystycznych genów niż we własnych dzieciach i wnukach.
Jednak Hamilton zwrócił uwagę na jeszcze jeden czynnik zwiększający interes genetyczny (który następnie rozwinęła genetyka populacyjna),mianowicie rzeczywisty status plemienia jako na wpół odizolowanej (za sprawą warunków geograficznych, znaczników kulturowych oraz warunków politycznych) populacji. W tej sytuacji pokrewieństwo winno być zobrazowane nie tyle jako drzewo rodowodowe, tylko jako sieć powiązań[1], którą nasi przodkowie intuicyjnie konceptualizowali jako rodzaj tkaniny, plecionej przez prządki losu – Rodzanice- splatające poszczególne linie rodowodowe wielokrotnie ze sobą na przestrzeni pokoleń, tworząc gęstą tkaninę wspólnego pokrewieństwa[2]. Hamilton tak właśnie konceptualizował interes genetyczny w swoim artykule z 1975 roku, gdzie w stosunku do plemienia/etnosu użył alegorii „miasta”. W jego modelu z powodu rosnącej sieci pokrewieństwa, wzrośnie ono między jednostkami do pewnego maksimum, wynoszącego 0,5, które odpowiada rodzeństwu w populacji panmiktycznej. Ten modelowy przykład jest tutaj kluczowy, gdyż ukazuje prawdopodobieństwo teoretyczne sytuacji w której etniczny interes genetyczny jest większy niż największe rodzinne dostosowanie łączne. Dla nas wnika z tego prosty wniosek, że altruizm ofiarny w imieniu etnosu może być adaptacyjny.
Populacje ludzkie na przestrzeni dziejów rozchodziły się w drzewie genealogicznym światowej populacji, populacje dzieliły się, a oddzielone fizycznie, różnicowały się genetycznie pod wpływem selekcji naturalnej, doboru płciowego, dryfu i mutacji. Jednocześnie wewnątrz, dzięki sieci wzajemnych powiązań krewniaczych i ograniczonej migracji utrzymywał się magazyn genetycznego interesu. Aby zobrazować ten stan F. Salter użył metafory wysokiego szczytu górskiego, który symbolizuje jednostkę, opadającego w dół tak jak jej pokrewieństwo z najbliższą rodziną, ale nagle przechodzącego w płaskowyż na poziomie etnosu, który tworzy grzbiet górski, przecięty uskokami między populacyjnymi. Takie gradienty wyznaczają granice etniczno- językowe nawet po wielu wiekach migracji i wojen. Dopiero te szerokie płaskowyże górskie, rozchodzą się w dół na inne kontynenty wraz z gwałtownym spadkiem pokrewieństwa. Patrząc z oddali na te łańcuchy górskie widać wyraźnie, że pod względem wysokości (symbolizującym pokrewieństwo) jednostka, jej najbliższa rodzina i etnos stanowi wyraźnie skoncentrowany obszar na dużej wysokości w stosunku do światowej populacji.
Jednak model ten, choć intuicyjnie logiczny, nie jest koncepcyjnie wyczerpujący w odniesieniu do zagadnienia pokrewieństwa wynikającego z badania ludzkich rodowodów, co czyni go podatnym na krytykę (jak ta ze strony Cosmides i Toobyego, opisywana w poprzedniej części eseju), zarówno merytoryczną jak i sofistyczną. [3] O wiele bardziej precyzyjne wyjaśnienie koncepcji dostosowania łącznego jak i GST, czyli odpowiedzi na pytanie dlaczego my, nacjonaliści, darzymy pozytywnym uczuciem pewne „obce” osoby i grupy, tymi samymi uczuciami co najbliższych krewnych, dzięki H. Harpendingowi dostarczyła genetyka populacyjna.
Przed opisaniem teorii etnicznego interesu genetycznego z perspektywy genetyki populacyjnej należy jednak zamieścić kilka zdań wprowadzających do zagadnienia.
Genetyka populacyjna zajmuje się, jak to wynika z nazwy, badaniem populacji, które są określane jako grupa organizmów należących do tego samego gatunku i żyjących w obrębie ograniczonego obszaru geograficznego.[4] Lokalne, wzajemnie krzyżujące się jednostki możliwie dużych populacji wykazujących strukturę geograficzną są obiektem zainteresowania, ponieważ to właśnie w takich jednostkach zachodzi ewolucja adaptacyjna w kierunku systematycznych zmian częstości alleli. Takie lokalne (ograniczone terytorialnie) wzajemnie krzyżujące się jednostki – zwane często populacjami lokalnymi lub demami – stanowią podstawowe jednostki genetyki populacyjnej. Lokalne populacje są w rzeczywistości ewoluującymi jednostkami gatunku[5].
Odkładając na bok zabawę słowną, zastosowaną przez autorów podręcznika, z którego została zaczerpnięta powyższa definicja genetyki populacyjnej (motywowaną zapewne polityczną poprawnością i strachem przed agresją i oskarżeniami o rasizm ze strony lewicowych aktywistów) możemy napisać wprost, że genetyka populacyjna zajmuje się procesami ewolucyjnymi na poziomie podgatunku, rasy i narodu/etnosu. Wracając do zagadnienia etnicznych interesów genetycznych, w przeciwieństwie do genealogii, genetyka populacyjna używa danych z testów genetycznych. W związku z powyższym zamiast kategorii stopnia pokrewieństwa, w tym przypadku mamy do czynienia ze współczynnikiem pokrewieństwa, który w istocie oznacza zmienność genetyczną lub dystans genetyczny. Trzecią charakterystyczną cechą tej perspektywy oglądu interesu genetycznego jest, jego względna natura, która wynika bezpośrednio z danych genetycznych, bez potrzeby stosowania obrazowych alegorii, jak to miało miejsce powyżej w przypadku stopnia pokrewieństwa. Jako, że zagadnienie to może być nieco skomplikowane dla czytelnika na co dzień nie obcującego z podobną tematyką, na końcu eseju został w całości zamieszczony tekst H. Harpendinga dot. współczynnika pokrewieństwa, do którego autor będzie się sukcesywnie odwoływał. Zatem omówmy po kolei za Frankiem Salterem wyszczególnione powyżej zagadnienia. Genetyka populacyjna odnosi się do współczynnika pokrewieństwa f, gdyż jest on precyzyjnie zdefiniowany matematycznie. Podstawowa różnica między tymi dwoma kategoriami zawiera się we wzorze (zob. załącznik):
r = 2f
Współczynnik pokrewieństwa f to prawdopodobieństwo tego, że gen znaleziony w genomie jednego osobnika w danym locus jest identyczny z genem znalezionym u innego osobnika w tym samym locus. Oznacza to, że gdy losowo pobieramy próbki genów od osób z różnych grup współczynnik pokrewieństwa jest miarą pokrewieństwa między grupami. Co charakterystyczne, w związku ze względną naturą pokrewieństwa, zawsze najważniejszy jest punkt odniesienia w postaci populacji bazowej (np. podgatunek) z której wyodrębnił się dany dem (etnos) lub po prostu średnie pokrewieństwo między ludźmi zawarte w światowej populacji. Ta nowa definicja pokrewieństwa pozwala takim porównywanym genetycznie parom osób mieć pokrewieństwo dodatnie, zerowe lub ujemne!
Jak pisze F. Salter globalny test genetyczny przeprowadzony w latach 80 ub. w przez L. Cavalli- Sforzę, P. Menozzi i A. Piazza mierzył nie pokrewieństwo, ale wariację lub inaczej „odległość genetyczną” wyrażaną jako współczynnik Fst. Na szczęście Harpending pokazuje, że pokrewieństwo można wyrazić kategoriach wariancji:
ƒo = Fst + (1 – Fst)[ -1/ (2N-1)]
gdzie ƒo jest lokalnym (demowym) współczynnikiem pokrewieństwa, Fst jest wariancją metapopulacji a N to cała populacja. Gdy N jest duże, jak to ma miejsce w przypadku porównywania współczesnych etnosów, dobrym przybliżeniem powyższego równania staje się po prostu:
ƒo = Fst
Oznacza to, że odległość genetyczna Fst jest zarówno miarą wariancji genetycznej (współczynnikiem pokrewieństwa) między dwiema populacjami, jak i miarą pokrewieństwa w każdej z nich[6]. Wynika z tego wprost relatywistyczna natura pokrewieństwa, która to ma kluczowe znaczenie dla dalszych rozważań na temat etnicznego interesu genetycznego.
Jak zatem zobrazować na prostych przykładach tę względną naturę współczynnika pokrewieństwa? Otóż na wstępie należy zauważyć, że słynne zdanie J.S. Haldana ilustrujące altruizm krewniaczy („Oddałbym życie za dwoje tonących braci lub ośmiu tonących kuzynów”) jest prawdziwe, jeżeli rodzice takich hipotetycznych krewnych byli by ze sobą spokrewnieni na poziomie zerowym, czyli gdy byliby losowo wybranymi osobami ze światowej populacji. Jeżeli jednak oboje rodziców będzie pochodziło z jednego etnosu ich pokrewieństwo nie będzie zerowe tylko dodatnie (odpowiadające średnio kuzynowi trzeciego stopnia). wtedy jak zauważył Ruschton w swojej teorii GST do połowy genów jednego rodzica dochodzi ok. 40% genów drugiego rodzica i pozostałe 10% genów które są wspólne dla obydwu rodziców[7]. W takim przypadku stężenia pokrewieństwa u potomstwa wzrastają powyżej 50%. Jest jednak druga strona tej względności. Jak pisze Salter „Co jeśli świat składałby się z kuzynów? Pokrewieństwo pomiędzy losowymi parami byłoby zerowe więc nie byłoby adaptacyjne dla nich okazywanie sobie altruizmu.”[8] W tym wariancie także słowa Haldana o ratowaniu ośmiu (lub nawet stu) kuzynów nie są poprawne, gdyż w świecie złożonym z samych kuzynów wyższa koncentracja pokrewieństwa występuje jedynie w rodzinie nuklearnej. W takim przypadku, adaptacyjnym byłoby okazywanie sobie altruizmu jedynie pomiędzy rodzicami i dziećmi oraz pomiędzy dziećmi, gdyż tylko tam pokrewieństwo byłoby czterokrotnie wyższe niż między kuzynami.
Jak konkluduje Salter, etnos jest analogiczny do populacji kuzynów. Gdyby światowa populacja składała się tylko z jednej grupy etnicznej, wtedy pokrewieństwo losowych rodaków byłoby (podobnie jak w przykładzie z kuzynami) zerowe. Zatem etniczne interesy genetyczne także byłyby zerowe wobec swoich rodaków. W tym wariancie altruizm byłby adaptacyjny jedynie wobec rodziny oraz dalszych krewnych.
W rzeczywistości wiemy doskonale, że biorąc pod uwagę populację całego świata pokrewieństwo losowych par wynosi zero. Gdyż jest ona wypadkową wielu różnych grup etnicznych. Wynika z tego wprost, że w przypadku dwóch losowych etnicznych Polaków, mówienie o nich jako o kuzynach etnicznych (lub tradycyjnie braciach ciotecznych) nie jest pozbawione realnych podstaw. Dane z testów genetyki populacyjnej, jak zobaczymy poniżej, wskazują, że łączne pokrewieństwo etniczne jest większe niż to zawarte w nawet największej rodzinie nuklearnej. Z drugiej strony odległość genetyczna między populacjami pochodzącymi z różnych kontynentów wzrasta raptownie o kilka rzędów wielkości.
Zanim przejdziemy do kwantyfikacji tego konceptu w odniesieniu do naszego etnosu i podgatunku, co z pewnością pomoże w zrozumieniu doniosłości powyższych ustaleń dla najbardziej żywotnych interesów naszego narodu i podgatunku, należy uczynić zadość popularno-naukowej rzetelności tego eseju. Aby nasz przegląd wiedzy naukowej był w pełni rzetelny należy pokrótce omówić pewne zastrzeżenia co do rzeczywistości etnicznych interesów genetycznych.
Podstawowe zastrzeżenie odnośnie etnicznych (i podgatunkowych) interesów genetycznych jest związane z upadkiem poglądu multiregionalnego lansowanego przez m.in. C. Coona, a dotyczącego rozprzestrzeniania się i różnicowania ludzi jako gatunku na ziemi, na rzecz koncepcji „out of Africa”.
Chociaż opisanie przebiegu i znaczenia debaty naukowej związanej z tym problemem wykracza poza ramy tej pracy, w pewnym uproszczeniu można stwierdzić, że niezależne potwierdzenie koncepcji „out of Africa” dostarczyło paliwa do pewnej sofistycznej krytyki teorii rzeczywistości podgatunkowej i rasowej oraz etnicznego interesu genetycznego[9]. Skoro bowiem, jak twierdzili krytycy koncepcji interesu genetycznego, początek światowej populacji ludzkiej zapewnili koloniści z Afryki którzy zaczęli migrować między 100 000 a 50 tysięcy lat temu na pozostałe kontynenty, to podczas tych migracji mieszali się oni ze sobą przez co nie można mówić o czystych liniach genealogicznych w odniesieniu do poszczególnych podgatunków tylko o jednej światowej populacji. Niewątpliwie przyczyniły się do tego nieprecyzyjne sformułowania twórcy drzewa filogenetycznego ludzkich populacji. L Cavalli Sforzy w odniesieniu do populacji europejskiej, która jego zdaniem zawierała istotne domieszki genów afrykańskich i azjatyckich ok. 30 000 lat temu.
W istocie, już na wstępnym etapie teoria genetycznego interesu etnicznego broni się, jeżeli weźmiemy pod uwagę niewielką Fst tych „afrykańskich” domieszek w stosunku do populacji europejskich. W istocie były to ludy tego samego pnia podgatunkowego, różniące się haplogrupą, które rozgałęziły się na kaukazie (stąd używane przez wielu badaczy określenie „rasa kaukaska” dotyczące metapopulacji) w trakcie zasiedlania subkontynentu europejskiego i śródziemnomorza. Podczas gdy haplogrupa „a „zasiedliła wcześniej północno – wschodnią Europę, tworząc od czasów społeczności Cro-Magnonskich, poprzez ostatnie zlodowacenie, społeczności łowców- zbieraczy, haplogrupa „b”pozostała częściowo w Azji Mniejszej i płn. Afryce, gdzie wzięła udział w rewolucji neolitycznej i dopiero po ustaniu zlodowacenia, u progu Holocenu zasiedlała w małych grupach, z południowego zachodu w kierunku północno - wschodnim Europę, powolnie mieszając się ze społecznościami łowców – zbieraczy, podlegając w czasie tej powolnej migracji tej samej presji selekcyjnej co wcześniejsze społeczności łowieckie.
Dodatkowo, jak zauważa Salter, sam Cavalli - Sforza twierdził, że tylko dryf genetyczny jest w stanie przez 40 000 lat od tego procesu skutecznie zróżnicować populację europejską od tych z innych kontynentów[10] Podobnie twierdził E. O. Wilson zakładając, że mikroewolucja populacji ludzkich może nastąpić w ciągu 1000 lat, główne adaptacje mogą potrwać 2000 lat a specjacja 40 000 lat. Prof K. Macdonald w swojej teorii zachodniego indywidualizmu prezentuje pogląd o następujących po sobie inwazjach indoeuropejskich wojowników ze stepów, którzy jednakowoż także stanowią pokrewną populację o dodatnim pokrewieństwie ze rdzenną ludnością europejską, co tłumaczy wg tego badacza błyskawiczną asymilację oraz niezmienne utrzymywanie się charakterystycznych dla europy cech fizycznych takich jak jasna pigmentacja skóry, oczu i włosów(nie występująca nigdzie indziej na świecie, mimo podobnych warunków ekologicznych) [11]
kolejnym dowodem odrzucającym twierdzenie że populacje europejskie są tak wymieszane, że pokrewieństwo etniczne jest praktycznie zerowe, są badania Briana Sykesa. Ten brytyjski genetyk po przebadaniu mitochondrialnego dna wskazał że tylko 15-20 % europejskiej puli genów pochodzi z Bliskiego Wschodu (kolebki rolnictwa). Regionalne skupiska mitochondrialnych DNA wskazały sześć populacji założycielskich, sięgających rodowodem 45 000 lat wstecz do czasów osad Cro- Magnon, a tylko jedna (siódma) to populacja rolników którzy weszli do Europy w neolicie. Jeżeli by w wyniku rewolucji neolitycznej i związanej z tym eksplozji demograficznej Europę zalały ludy z bliskiego wschodu, Europejczycy po prostu nie mogliby wyśledzić swoich przodków aż do Cro-Magnonczyków, którzy 45 000 lat temu wytrzebili Neandertalczyków.
Na koniec przypomnijmy że dla genetycznej specyfiki rdzennej populacji europejskiej, ogromne znaczenie miały zjawiska nazwane Czarnymi Łabędziami[12], których nagłe pojawienie się było selekcyjnym „wąskim gardłem” ewolucji. Zdarzenia takie jak epidemie chorób, czy nagłe ochłodzenia klimatu jeszcze bardziej przyspieszały tworzenie się unikalnych adaptacji będących kolejnymi pokładami etnicznych i podgatunkowych interesów genetycznych europejskich populacji.
Powyższe trzy koncepcje naukowe w innych okolicznościach historycznych mogłyby być jedynie interesującym elementem pewnej naukowej debaty, elementem akademickich dyskusji w której równorzędną rolę mogłaby odgrywać również lewicowa perspektywa wraz ze swoim biologicznym negacjonizmem. Jednak obecnie, po ponad pół wieku totalnej dominacji lewicy w europejskiej nauce, polityce i kulturze, destrukcyjny wpływ jej postulatów doprowadził nasze populacje na skraj egzystencjalnej przepaści. Mimo to lewica wciąż brnie naprzód w swoich dysgenicznych postulatach, doprowadzając je do granicy absurdu (a czasem nawet ją przekraczając) co gorsza w ślad za nią podąża część nacjonalistów, mniej lub bardziej świadomie biorąc udział w dziele zniszczenia naszej rodzimej tradycji oraz genetycznych interesów. W epoce masowych migracji skutkujących wymianą rdzennej ludności europejskiej, etniczne interesy genetyczne indoeuropejskich narodów są zagrożone całkowitym zniszczeniem. Problem polega głównie na tym, że nawet gwałtowny spadek liczebności populacji może być całkowicie niedostrzegalny z perspektywy jednostki lub rodziny nuklearnej, podczas gdy śmierć dziecka w takiej rodzinie jest szokiem i tragedią, a nawet śmierć osoby starej we śnie (lub z powodu korona- grypy) oznacza żałobę w każdej rodzinie. Tymczasem w rzeczywistych wartościach, spadek liczebny populacji oznacza większy ubytek genetycznych odsetek każdego członka narodu, niż śmierć kogokolwiek z jego rodziny. Jednak, jak to zostało powiedziane powyżej, ubytek interesu genetycznego nie musi wynikać z prostego wymierania. Pewne procesy są powiązane z państwem, które jest polityczną i terytorialną organizacją etnosu. Państwo działające w sposób adaptacyjny koordynuje wytwarzanie dóbr publicznych takich jak instytucje publicznej socjalizacji, służba zdrowia, straż pożarna itp. oraz spełnia swoją podstawową funkcję jaką jest ochrona terytorium. Ochrona terytorium jest żywotnym interesem państwa, gdyż zapewnia ochronę interesu genetycznego etnosu. Ułatwia ona ciągłość puli genów etnosu, zwiększając zdolność grupy etnicznej do obrony przed masową imigracją zarówno agresywną jak i pokojową [13].
Na przekór demoliberalnym piewcom „końca historii” obecna europejska rzeczywistość wyraźnie wskazuje, że obrona terytorium wciąż jest podstawową strategią grupy etnicznej - jest ona obecnie równie adaptacyjna jak w pierwotnych środowiskach - podczas kryzysu imigranckiego lub pandemii tylko bronione granice zapewniają państwu skuteczną reakcję na zagrożenie. Zdziesiątkowana, pokonana lub zubożała populacja ma szansę na regenerację, gdy zachowa kontrolę nad swoim terytorium. W przeciwnym razie napływ na dużą skalę genetycznie odległych imigrantów nieodwracalnie zmniejszy interes genetyczny rdzennej populacji.
W tym miejscu pojawia się pytanie dlaczego ten proces będzie nieodwracalny? Odpowiedź na to zagadnienie można znaleźć w ustaleniach naukowych z zakresu ekologii. Dwóch badaczy zajmujących się tym obszarem wiedzy poczyniło ustalenia składające się na koncepcję nośności ekologicznej danego terytorium oraz całej planety. Pierwszy z nich, G. Hardin, niestrudzony krytyk liberalizmu z jego koczowniczą mentalnością „pastwiska” zwrócił uwagę, na rażący brak cech adaptacyjnych kapitalistycznego stosunku do ekosystemu[14]. Nośność ekologiczna zarówno poszczególnych obszarów, jak i całej planety jest ograniczona. Jeżeli na danym obszarze zostanie przekroczona jego nośność poprzez wzrost populacji lub wykorzystywanie jego zasobów (albo jedno i drugie) nastąpi ograniczenie liczby ludności poprzez wymieranie spowodowane brakiem zasobów. Dotychczas postęp technologiczny zwiększał nośność terytorium ale i on zdaje się wyczerpywać. Drugim badaczem który zwrócił uwagę na tego typu problemy jest doskonale znany z poprzednich tekstów E.O. Wilson, który w pracy „Pół Ziemi” stworzył koncepcję „śladu ekologicznego”. Identycznie jak Hardin doszedł on do wniosku, że planeta jest przepełniona i aby zatrzymać masowe wymieranie całych ekosystemów, należy ograniczyć ów „ślad ekologiczny”(w którego skład wchodzą średnia ilość ziemi produkcyjnej i płytkiego morza (szelfu) przypadająca w kawałku na każdą osobę, z którego to czerpie ona żywność, [15] wodę, mieszkanie, energię, transport, handel i absorbcję odpadów).
Na marginesie widać tu kolejną absurdalność lewicowej ekologii, która mając na sztandarach ochronę ekosystemów, jednocześnie promuje imigrację z obszarów biednego południa globu, do krajów uprzemysłowionej północy. Nietrudno się domyśleć że dla całej planety takie zachowanie będzie miało katastrofalne skutki. O ile bowiem w krajach rozwijających się, na jednego mieszkańca przypada około jednego hektara śladu ekologicznego, to na mieszkańca USA ślad ekologiczny wynosi 9,6ha – gdyby więc przenieść całą ludność świata do USA i utrzymać tamtejszą konsumpcję na obecnym poziome, potrzebne byłyby cztery kolejne planety wielkości Ziemi!
Zdaniem Hardina populacje naszej planety muszą ograniczyć dalszy wzrost (do poziomu 2,1 czyli zastępowalności pokoleń) gdyż ślad ekologiczny populacji nie może w dłuższej perspektywie przekraczać terytorium które ona kontroluje.
W tym właśnie zawiera się odpowiedź na kwestię nieodwracalności procesu utraty etnicznego interesu genetycznego. Jeżeli bowiem europejskie społeczeństwa (a także płn.wschodnio azjatyckie) są w stanie narzucić sobie taką dyscyplinę, stają się narażone na imigrację. Jeżeli etnos, kontrolujący dane terytorium, za powszechnym porozumieniem postanawia chronić własny ekosystem nie przekraczając nośności ekologicznej przewyższającej rozwój technologii zmniejszających ślad ekologiczny, to każda imigracja która nie niesie ze sobą takiego właśnie postępu technologicznego, zmniejszy wielkość rodzimej populacji [16]. Chociaż Hardin nie bierze pod uwagę etnicznych interesów genetycznych, tylko indywidualne dostosowanie wnioski są oczywiste: każde 10 000 imigrantów obniży efektywną nośność ekologiczną kraju o 10 000 (zakładając dla uproszczenia, że imigranci i rdzenni mieszkańcy mają tą samą dzietność). Jednak prawdziwy rozmiar problemu można dostrzec jedynie wtedy, gdy w tym modelu uwzględnimy koncepcję etnicznego interesu genetycznego. Wtedy, w oparciu o wcześniejsze ustalenia, w zależności czy imigranci przybywają z pokrewnych środowisk etnicznych, rdzenna populacja nie doznaje znaczącego uszczerbku interesu genetycznego (bliżsi krewni są zastępowani dalszymi krewnymi). Jeżeli jednak mamy do czynienia z wymianą ludności ze środowisk genetycznie odległych, nastąpi ogromna utrata etnicznego interesu genetycznego. Aby to zagadnienie zobrazować posłużmy się kwantyfikacją interesu genetycznego dokonaną w oparciu o metody wypracowane przez genetykę populacyjną.

Tabela 1. odległości Fst pomnożone przez 10 000 pomiędzy wybranymi populacjami na całym świecie (zaczerpnięte z Cavalli- Sforza i inni 1994 s,75)
Przeglądając powyższą tabelę F. Salter zwraca naszą uwagę na pozorny paradoks wynikający ze względnej natury genetycznego pokrewieństwa (współczynnik pokrewieństwa f). Współczynniki pokrewieństwa wynoszą odpowiednio 0,0021 w przypadku blisko spokrewnionych genetycznie Anglików i Duńczyków na jednym końcu skali, do 0,43 w przypadku najbardziej odległych genetycznie od siebie australijskich Aborygenów i afrykańskich Pigmejów Mbuti. Dlaczego tak się dzieje skoro w drzewie genealogicznym europejskich populacji Anglicy są tuż obok Duńczyków, natomiast w drzewie genealogicznym o zasięgu światowym Mbuti są na jednym skraju a Aborygeni na drugim? Ten pozorny paradoks wyjaśnia się, gdy powrócimy do definicji pokrewieństwa genetycznego i jego natury. Jak pamiętamy pokrewieństwo genetyczne mierzy zarówno pokrewieństwo w obrębie populacji jak i, z odwrotną zależnością, pomiędzy populacjami. Tak więc tutaj sytuacja między Anglikami i Duńczykami jest analogiczna do naszego modelu teoretycznego gdzie światowa populacja składała się z samych kuzynów. Jest tak właśnie dlatego, że Anglicy i Duńczycy są blisko spokrewnieni. Gdyby świat składał się z samych Anglików pokrewieństwo pomiędzy losowymi rodakami byłoby zerowe. Ale w świecie złożonym z Anglików i Duńczyków pokrewieństwo losowego Anglika z innym Anglikiem wzrasta nieznacznie o 0,0021 (odpowiednik pradziadka w szóstym pokoleniu). Innymi słowy rdzenny Anglik nie straciłby wielu swoich charakterystycznych genów, gdyby jego rodak został zastąpiony Duńczykiem.
W przypadku losowego Aborygena i Pigmeja, ze względu na odwrotną relację między pokrewieństwem wewnątrzetnicznym a miedzyetnicznym, losowy Aborygen traci o wiele więcej genów jeżeli jego rodak zostanie zastąpiony przez Pigmeja z Afryki, który jest bardzo odległy genetycznie. W tym przypadku, w świecie złożonym z samych Aborygenów i Pigmejów, pokrewieństwo genetyczne dwóch losowych Aborygenów wzrasta do 0,43 co oznacza prawie pokrewieństwo z bliźniakiem monozygotycznym! (lub z samym sobą – patrz załącznik). Oznacza to innymi słowy, że pojedyncza migracja Pigmeja na teren Aborygenów oznacza dla każdego z nich utratę prawie bliźniaka jednojajowego.
Przy założeniu ograniczeń ekologicznych związanych z nośnością ekologiczną każdego etnicznego terytorium opisywanymi powyżej, współczynniki Fst opracowane przez Cavalli- Sforzę ukazują,że imigracja jest tym bardziej szkodliwa dla interesów genetycznych populacji rodzimej, im bardziej genetycznie odlegli są imigranci. Aby jeszcze bardziej zobrazować koncepcję utraty etnicznego interesu genetycznego, poniesionej w wyniku imigracji krzyżowej pomiędzy populacjami umieszczonymi w tabeli 1 można tę stratę zamienić na ekwiwalent dziecka. Jako, że w prawie każdej ludzkiej kulturze strata dziecka jest największą ludzką tragedią, pojawiający się powyżej ekwiwalent w postaci bliźniaka, zmieńmy na ekwiwalent dziecka, tym bardziej, że H. Harpening (w załączniku na końcu artykułu) wyprowadził ze współczynnika Fst wzór na współczynnik pokrewieństwa z własnym dzieckiem. Będzie to:
ƒ= 0,25 + ¾ Fst
Jeżeli podstawimy do pow. wzoru najbliżej spokrewnionych genetycznie Anglików i Duńczyków ze wzajemnym Fst=0,0021 i założymy, że imigracja będzie polegała na zastąpieniu 10 000 Anglików przez 10 000 Duńczyków przy innych zmiennych na tym samym poziomie (współczynnik dzietności, śmiertelność itp.) Pierwszą stratą interesu genetycznego będzie więc usunięcie 10 000 Anglików co daje nam 0,0021 x 10 000 czyli -21 jednostek pokrewieństwa, a następnie kolejne – 21 jednostek pokrewieństwa spowodowane podmienieniem w to miejsce 10 000 Duńczyków co daje kolejne -21 jednostek pokrewieństwa czyli razem mamy utratę 42 jednostek angielskich genetycznych interesów. Aby następnie tę wielkość zobrazować liczbą straconych dzieci musimy ją podzielić przez pokrewieństwo rodzica z dzieckiem wewnątrz populacji angielskiej czyli:
ƒ= 0,25 + (0,0021 x ¾) = 0,2516
zatem liczba utraconych dzieci w wyniku migracji 10000 obcych o Fst 0,0021 wynosi
42/0,2516 ≈ 167
Każdy kto w tym momencie myśli, że strata 167 dzieci angielskich w wyniku migracji 10 000 Duńczyków jest dużym problemem, powinien zatem przeliczyć stratę wywołaną imigracją i zastąpieniem rodzimej populacji Anglików dziesięcioma tysiącami Subsaharyjczyków z plemienia Bantu. W tym przypadku mamy:
10 000 x 0,2288=- 2288 straty wywołanej utratą Anglików, oraz drugą taką stratę powstałą, gdy miejsce utraconych 10 000 rodowitych Anglików zostanie zajęte przez 10 000 Bantu, czyli razem:
-2288-2288 = 4576
oraz pokrewieństwo rodzica z dzieckiem w populacji angielskiej wobec Fst pomiędzy losowym Anglikiem a Bantu:
ƒ= 0,25 + (0,2288 x ¾) = 0,1716 + 0, 25 = 0,4216
Zatem w tym przypadku, zsumowanie straty Anglików wynoszące 4576 dzielimy przez pokrewieństwo Anglika z dzieckiem czyli 0,4216 co daje
4576 / 0,4216 ≈ 10,854
utraconych dzieci.
Dlaczego jednak w tym przypadku liczba utraconych dzieci jest wyższa niż liczba imigrantów zastępujących rodzimą populację angielską? Salter tłumaczy to następująco; W rzeczywistości tak nie jest, gdyż liczymy geny a nie osoby. Wynik wygląda dziwnie tylko wtedy gdy liczba genów jest konwertowana na odpowiedniki podrzędne, chociaż konwersja jest prawidłowa. Losowi członkowie grupy etnicznej są skoncentrowanymi magazynami charakterystycznych dla siebie genów, podobnie jak ich kuzyni i dzieci. Niektóre grupy etniczne są tak różne od siebie genetycznie, że stanowią ujemne zapasy tych charakterystycznych dla siebie genów. Imigracja [obcych podgatunkowo populacji] ma więc podwójny wpływ na dostosowanie. Po pierwsze zmniejsza potencjalny pułap populacji rodzimej i po drugie, trwale zastępuje charakterystyczne geny utraconych rodaków obcymi, egzotycznymi odmianami[17]. Wszystko więc zależy od stopnia pokrewieństwa między populacjami. Tam gdzie jest ono duże, straty wywołane imigracją są niewielkie, tam gdzie jest duża genetyczna zmienność, straty w kontekście ekologicznego pułapu terytorium mogą być nieodwracalne. Kiedy bowiem naród w wyniku wojny lub epidemii traci nawet znaczną część populacji ale jest zabezpieczony granicami przez imigracją, zawsze z czasem zregeneruje się liczebnie (a więc także w wymiarze etnicznego interesu genetycznego), do pułapu wyznaczonego przez nośność ekologiczną swojego terytorium. Natomiast zajęcie przez obcy etnos luki wywołanej utratą części populacji oznacza, że nie urodzą się konkretne dzieci i wnuki, gdyż zostaną zastąpione obcymi o ujemnym stężeniu rodzimych genów. Tutaj zwycięzcą będą imigranci, gdyż ich ubytek w ojczyźnie zostanie zastąpiony rodakami, zwiększając genetyczny interes populacji.
Jak wynika z powyższych szacunków, altruizm etniczny włącznie z ofiarnym heroizmem jest tym bardziej adaptacyjny, im bardziej wspiera rodaków etnicznych w rywalizacji lub walce z etnicznie odległymi imigrantami bądź najeźdźcami, szczególnie jeżeli pochodzą oni z innych ras geograficznych.Rysunek nr.2 ukazuje drzewo genealogiczne genetycznych klastrów ludzkich (o których pisałem w poprzednim eseju), natomiast tabela 2 ukazuje miary odległości genetycznej pomiędzy tymi klastrami. Podobnie jak inne różnice między tymi populacjami (określone jako rasowe) dystans genetyczny pomiędzy Anglikami i Bantu jest tak duży, że konkurencja między nimi spowodowana imigracją, sprawiłaby, że altruizm wobec przypadkowego rodaka równa się altruizmowi wobec własnego dziecka. Jak pisze Salter (parafrazując przywołanego wcześniej Haldane’a) bardziej adaptacyjnym wydaje się dla rdzennego Anglika ryzykowanie życia, mienia lub wolności w celu oporu przeciwko imigracji do Anglii dwóch Bantu niż podejmowanie takiego samego ryzyka w celu uratowania jednego z własnych dzieci przez utonięciem.
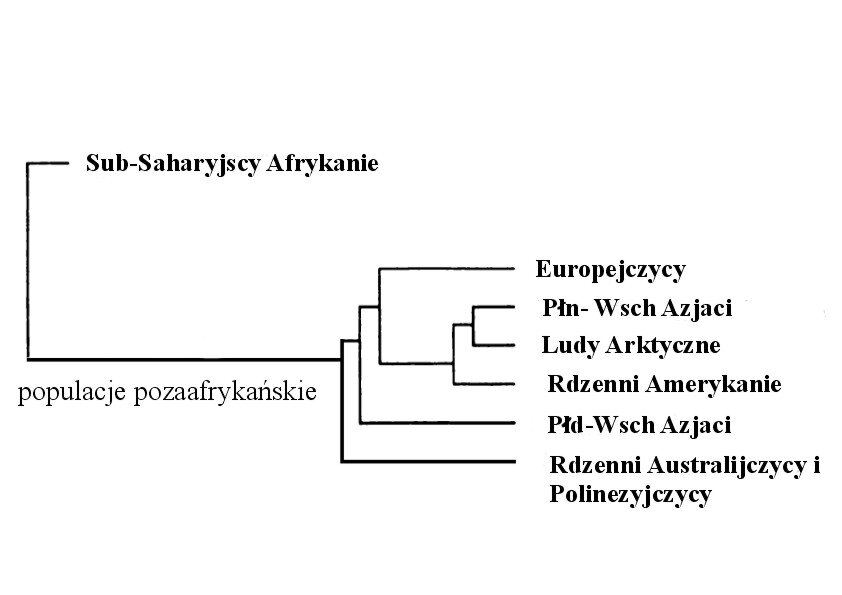
Rysunek 2 Odległości genetyczne pomiędzy siedmioma geograficznymi rasami.(Spisane z Cavalli-Sforza i in 1994 s. 79)
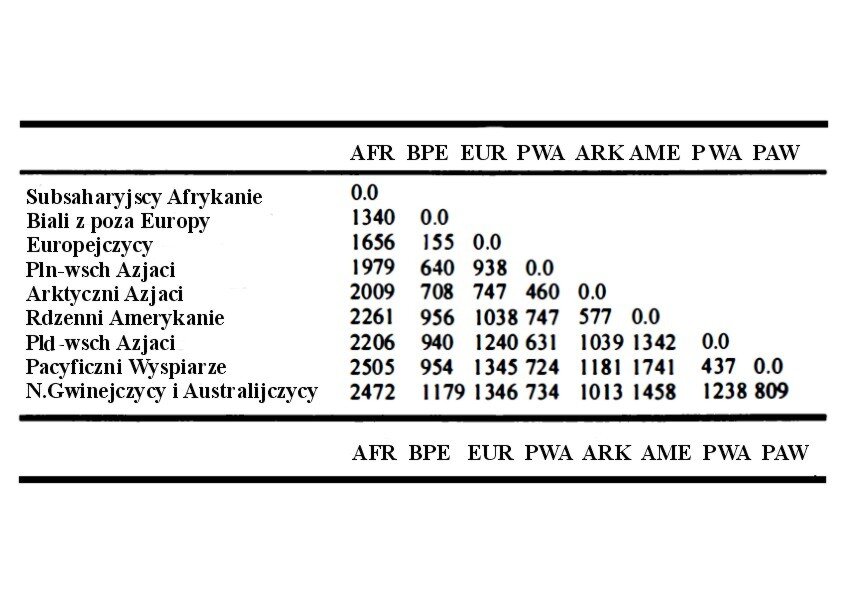
Tabela 2. Współczynniki pokrewieństwa rasowego dziewięciu ras geograficznych x 10 000. Rasowe pokrewieństwo między losowymi osobami różni się w zależności od rodzaju rywalizacji (np. Potencjalna imigracja) między różnymi rasami. Przypadkowi współplemieńcy mają zerowe pokrewieństwo, gdy grupa etniczna jest rozpatrywana w wariancie modelowym opisanym powyżej gdy świat składa się tylko z tej grupy. (Na podstawie zmienności Fst podanych przez Cavalli-Sforza i in. 1994, s. 80: zaokrąglone do najbliższych liczb całkowitych; pominięto standardowe błędy).
Międzypodgatunkowa imigracja jest najbardziej szkodliwa dla etnicznych interesów genetycznych, bez względu na pożądany przez obywatelskich nacjonalistów stopień asymilacji. Staje się to oczywiste, jeżeli weźmie się pod uwagę liczbę imigrantów potrzebną do zmniejszenia etnicznego interesu genetycznego losowego tubylca poprzez zamianę jego dziecka na imigranckie (przy tych samych pozostałych warunkach). Chyba najbardziej wymowne będzie porównanie tabeli 3 i 4 które pokazują kolejno liczby zastępcze dla imigracji międzyrasowej i liczby imigrantów w zasadniczo jednorodnym etnicznie kontynencie europejskim.
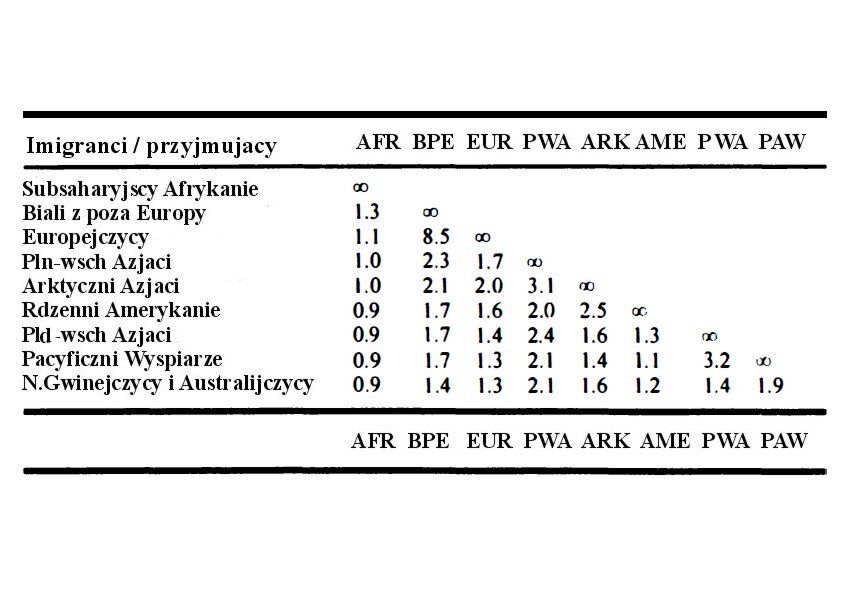
Tabela 3. Liczba imigrantów między dziewięcioma rasami geograficznymi potrzebna do zmniejszenia etnicznego interesu genetycznego przypadkowego tubylca o równowartość jednego dziecka

Tabela 4. Liczba imigrantów między 26 europejskimi etnosami potrzebna do zmniejszenia etnicznych interesów genetycznych przypadkowego tubylca o równowartość jednego dziecka. (Na podstawie odległości genetycznych Fst podanych przez Cavalli-Sforza i wsp., 1994, s. 270).
Widać wyraźnie, że imigracja obcych podgatunkowo populacji do Europy, skutkująca zastąpieniem przez nie rdzennych etnosów jest o kilka rzędów wielkości gorsza dla genetycznych interesów niż imigracja wewnątrzeuropejska ( próg jest przeważnie od 10 do 100 razy wyższy niż imigracja obca podgatunkowo). Kontrast ten jest uderzający bez względu na jakiekolwiek wymówki obywatelskich Bąkiewiczów. Jeżeli dla obniżenia średniego europejskiego interesu genetycznego o równowartość jednego dziecka wystarczy jedynie 1,1 subsaharyjskiego imigranta (patrz tabela 3) to naszych rodaków potrzeba aż 27 aby obniżyć interes genetyczny Niemców o równowartość 1 dziecka. Kiedy zaś spojrzymy na najbardziej podobnych genetycznie do naszego narodu Węgrów oraz etnicznych Rosjan to potrzeba ich odpowiednio 50,4 i 42 aby obniżyć nasz interes genetyczny o ekwiwalent dziecka.
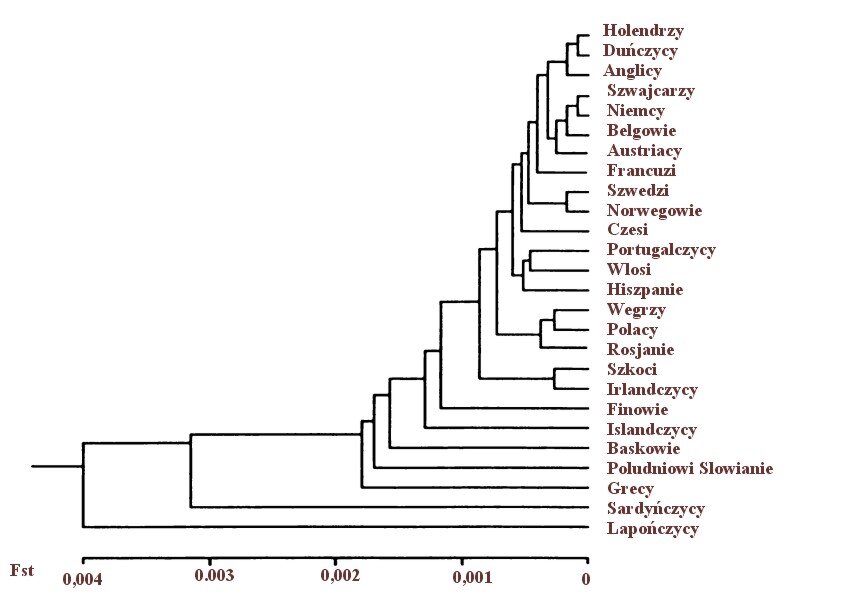
Rycina 3 Drzewo genetyczne 26 populacji europejskich w oparciu o oszacowane miary Fst z testu 88 genów (z Cavalli-Sforza i wsp. 1994, str. 268).
Jak widać, powyższe dane są najbardziej fundamentalnym uzasadnieniem etnicznego nacjonalizmu co ważne, nie chodzi tutaj o wartościowanie poszczególnych etnosów na podstawie arbitralnych kategorii kulturowych, ekonomicznych czy religijnych. Dane genetyczne z testów Fst nałożone na mapę światowych etnicznych migracji skutkujących podmianą populacji [18] są po prostu predyktorem egzystencjalnych problemów każdego kraju, którego dotyka to zjawisko.
Każdy kto deklaruje się jako obywatelski nacjonalista powinien zatem jasno sprecyzować dlaczego, programowo dopuszczając imigrację do Polski obcych etnosów rujnujących przyszłość kolejnych generacji naszych rodaków bierze w tym udział. Różnorodna masa badań dotycząca populacji dotkniętych multikulturalizmem nie pozostawia bowiem wątpliwości co do dalekosiężnych efektów utraty genetycznego interesu populacji. Zresztą, w dobie narastającej rywalizacji hegemonicznej pomiędzy USA a Chrld coraz bardziej uwidacznia się zadyszka tego pierwszego mocarstwa (mającego teoretycznie wszystkie atuty po swojej stronie), wobec tempa wyzwań rzucanych mu nieustannie przez homogeniczne i etnocentryczne Chrld. Równie dobrze uwidacznia się to w coraz poważniejszym i pernanentniejszym kryzysie ekonomicznym wywołanym przez finanse, pasożytnicze koncerny oraz giełdowych spekulacyjnych free-riderów. Najbardziej jednak znamienny jest obecny kryzys związany z tzw. pandemią korona-grypy, w którym, poza wszystkimi innymi aspektami tego zjawiska potwierdzenie znajdują badania Vanhanena (2012) twierdzącego, że jeśli społeczność wieloetniczna stanie w obliczu zagrożenia z zewnątrz, będzie się zachowywać w mniej etnocentryczny sposób niż populacja monoetniczna [19]. To samo wykazywał belgijski socjolog Marc Hoghe (2009) który, wraz ze współpracownikami wykazał związek między różnorodnością etniczną a spójnością społeczną, zwłaszcza zaufaniem [20]. Nowsze badania z Wielkiej Brytanii potwierdzają wcześniejsze ustalenia (Sturgis i inni 2011) choć badacze podkreślają, że wielkość efektu jest niewielka ale znacząca”. Także badania innego Brytyjczyka (James Laurence 2008) potwierdzały, że spójność społeczności GB jest zmniejszona, im bardziej zróżnicowana etnicznie jest społeczność. Co ciekawe w kontekście bieżących wydarzeń (epidemia) ten sam badacz w późniejszych badaniach stwierdza, że różnorodność podważa kapitał społeczny na poziomie sąsiedztwa, ale nie zmniejsza ogólnego poziomu indywidualnego zaangażowania. Oznacza to, że ludzie przestają uczestniczyć w działaniach społeczności i po prostu wykorzystują ten czas, aby spędzić go z rodziną i przyjaciółmi – czyli tymi, którzy są do nich bardziej genetycznie podobni niż szersza społeczność krajowa. To odkrycie, choć nie rozwinięte przez Laurence'a, jest interesujące, ponieważ implikuje, że życie w zróżnicowanym etnicznie obszarze zmniejsza ważny aspekt etnocentryzmu na poziomie grupy, ale może nawet zwiększyć go na bliższym poziomie genetycznym [21]. Tłumaczyłoby to dlaczego w dobie koronawirusa ludzie są tak przejęci śmiercią bliskich w podeszłym wieku (których odejście w wyniku choroby i tak już było przesądzone i nie miało wpływu na genetyczne interesy) jednocześnie lekceważąc destrukcyjne skutki ograniczeń sanitarnych na krajową gospodarkę.
Utrata interesu genetycznego spowodowana imigracją będzie miała również negatywny wpływ na kulturę, religię i tradycję rodzimej populacji, której obronę tak mocno akcentują Obywatelscy nacjonaliści. Jak wykazał prof. K. Macdonald podstawową strategią grup mniejszościowych zapewniającą im sukces w rywalizacji z grupą większościową jest właśnie niszczenie kluczowych adaptacji (Religia i tradycja) zapewniających spójność społeczną (żywotność społecznej tkanki). Z drugiej strony w oparciu o podane wyżej ustalenia Hamiltona i dane z genetyki populacyjnej można przewidzieć, że populacja wielokulturowa będzie miała jako całość bardzo niski pozytywny etnocentryzm (objawiający się brakiem gotowości do poświęcenia zasobów za całość społeczeństwa przy jednoczesnym zwiększonym negatywnym etnocentryźmie, objawiającym się wzajemną wrogością wewnętrznych grup wobec siebie, co wynika ze względnej natury pokrewieństwa genetycznego. Tak więc, jak twierdzi Vanhanen im większe są grupy etniczne wewnątrz populacji, tym większa wrogość między nimi skutkująca wzrostem heterogeniczności kraju (państwa) jako całości. Kraj taki stanie się areną wewnętrznego konfliktu rasowego, którego granica będzie przebiegać wzdłuż genetycznych gradientów między populacjami. Na podstawie wielkości mniejszości rasowej Vanhanen stworzył miarę [22] heterogeniczności etnicznej, będącej predyktorem występowania w tym kraju konfliktów o różnym natężeniu. W oparciu o swoją miarę Vanhanen przebadał 176 krajów stwierdzając, że istnieje korelacja na poziomie 0,66 pomiędzy rozmiarem konfliktu etnicznego a poziomem heterogeniczności etnicznej. Jednocześnie z drugiej strony istnieje negatywny związek pomiędzy heterogenicznością [23] (konfliktem etnicznym) a standardem życia.
Oczywistym pytaniem, które w związku z zawartą tu miażdżąca krytyką obywatelskiego nacjonalizmu jako ideologii obniżającej homogeniczność populacji, zadaje sobie każdy czytający te słowa podkomendny Bąkiewicza lub innych genetycznych mutantów brzmi: Dlaczego podgatunkowa, rasowa i etniczna tożsamość (której naukowy fundament został opisany w tym eseju), która jest mi droga i którą instynktownie akceptuję, jest moralnie potępiana przez funkcjonariuszy pustynnej religii, którą wyznaję? Ta kwestia z pewnością jest kluczowa jeżeli chcemy skutecznie odpowiedzieć na krytykę genetycznego interesu etnicznego sformułowaną z pozycji moralnych, wysnutych z tzw. nauczania kościoła.
Krytyka etnicznego interesu genetycznego, oraz etnocentryzmu jako takiego z pozycji katolickich może przybierać dwie formy.
Po pierwsze będzie to krytyka etnicznego nacjonalizmu jako strategii ewolucyjnej sprzeciwiającej się imigracji etnosów i grup religijnych jawnie wrogich zachodniej cywilizacji. W pierwszej kolejności chodzi tu o krytykę ze strony postsoborowego kościoła skierowaną wobec tzw. islamofobii [24] oraz nacjonalistów, wyrażających sprzeciw wobec mniejszości, realizujących własne strategie ewolucyjne kosztem rodzimego etnosu. Oczywiście krytyka ta, nie dysponując żadnymi argumentami, mającymi praktyczne odniesienie w rzeczywistości, kiedy wyczerpie się argumentacja zaczerpnięta z lewicowej nowomowy polegająca na mówieniu o „ubogacaniu”, „urozmaicaniu” itp. zawsze na końcu odwołuje się do Nowego Testamentu, przywołując wersy z pism Mateusza (5,44) oraz Łukasza (6,27) „Miłujcie nieprzyjacioły wasze”.
Zdając sobie sprawę z wiążącej mocy „słowa bożego” dla każdego katolika, piszący te słowa nie ma zamiaru go podważać, tym bardziej, że dla obrony wobec krytyki etnicznego interesu genetycznego nie jest to wcale konieczne. Dzieje się tak, dzięki dorobkowi intelektualnemu najmłodszego klasyka myśli prawniczej[25], wybitnego historiozofa i politologa Carla Schmitta. W swojej pracy „Pojęcie polityczności” niemiecki jurysta rozbraja całkowicie ten religijny argument, dogłębnie wskazując i opisując nie istniejący w większości języków indoeuropejskich rozdział znaczeniowy pomiędzy wyrazami „wróg” i „nieprzyjaciel” (rozróżnienie pomiędzy wrogiem publicznym a prywatnym) pisząc: Wróg nie jest więc konkurentem ani przeciwnikiem w sensie ogólnym. Wróg nie jest też prywatnym przeciwnikiem, którego nienawidzimy czy do którego czujemy osobistą antypatię. Wróg to przynajmniej potencjalnie tj. w sensie realnej możliwości walcząca grupa ludzi stojąca naprzeciw podobnej grupy. Wróg ma charakter wyłącznie publiczny, wszystko bowiem co odnosi się do takiej grupy ludzi, a w szczególności do narodu ma sens publiczny. Wróg to hostis a nie inimicus o szerszym znaczeniu […] Często przytaczane zdanie „miłujcie nieprzyjacioły wasze”b rzmi „diligite ininimicos vestros” nie zaś „diligite hostes vestros”. O wrogu politycznym nie ma zatem mowy. W tysiącletniej historii wojen między chrześcijaństwem a islamem nigdy żaden chrześcijanin nie wpadł na pomysł aby z miłości do Saracenów lub Turków wydać im na łup Europę, zamiast stanąć w jej obronie. Politycznego wroga nie trzeba wcale osobiście nienawidzić. Postulat miłowania „wroga” ma sens dopiero w sferze prywatnej. Przytoczony fragment Biblii nie zawiera zniesienia różnicy między dobrem a złem, ani między pojęciami piękna i brzydoty, a już w najmniejszym stopniu nie zachęca do porzucenia różnicy między przyjacielem a wrogiem. W każdym razie nie zaleca, aby kochać wrogów swojego narodu i wspierać ich w walce przeciwko własnym rodakom[26]
Drugi rodzaj krytyki ze strony kościoła katolickiego, skierowanej wobec etnicznego interesu genetycznego wynika z analogicznego jak w przypadku obywatelskich nacjonalistów, przejęcia przez postsoborowy kościół, oświeceniowej aksjologii. Ten ideologiczny nowotwór, rozwinął się na organizmie kościelnej wspólnoty do tego stopnia że nawet osoby będące formalnie poza jej głównym ideologicznym nurtem jak np. dr. E. Michael Jones [27] za fundament własnego stanowiska wobec genezy narodów, przyjmują quasi boasowską koncepcję religii jako genezy i źródła tożsamości etnosu. Dr Jones, polemizując z etnicznym i rasowym patriotyzmem w USA stwierdza, że Indoeuropejczycy w tym kraju przyjęli białą tożsamość tylko dlatego, że pozbawiono ich prawdziwej tożsamości (tej religijnej) dochodząc do tego na podstawie Heglowskich „Wykładów z filozofii dziejów”. Mamy więc tutaj stanowisko oparte na oświeceniowym idealizmie filozoficznym – narody stworzyła religia, lecz jest to tylko etap przejściowy na drodze postępu, którego zwieńczeniem będzie idealnie harmonijna ludzkość zjednoczona duchowymi ideałami chrześcijaństwa. Dodajmy, że pisze to katolicki radykał uchodzący za „świętszego od papieża”. Jones oczernia tożsamość rasową jako materialistyczną i sztuczną, całkowicie ignorując fakt, że nawet jeżeli, jako biali nie jesteśmy zainteresowani naszą rasową tożsamością to i tak według tej właśnie kategorii postrzegają nas nasi wrogowie, co czyni z niej niezdejmowalny mundur. Inną sprawą jest typowo lewicowa maniera Jonesa, aby nasze stanowisko przedstawiać jako skrajny determinizm genetyczny, całkowicie pomijając jego kulturowo - genetyczny aspekt. Ostatnia praca pt. „Etnos need Logos” zamiast być argumentem przeciw etniczności, jest w istocie ponurym świadectwem upadku intelektualnego katolicyzmu, który został ideologicznie skolonizowany przez lewicowe prądy oświecenia. Jones pisząc Rasa […] jest dziełem materializmu biologicznego, który znalazł swojego najwybitniejszego rzecznika w Karolu Darwinie. Materializm opiera się na prymacie materii a materia jak wie każdy wyszkolony z filozofii tomistycznej jest zasadą różnicowania. Materia nie może zatem prowadzić do jedności [28]uznaje tożsamość rasową za grzech, po lewacku ignorując rzeczywistość relacji między rasowych w imię utopijnego ideału harmonijnej ludzkości, zjednoczonej przez duchowe wartości. Co gorsza, próbuje angażować w to intelektualny dorobek naszej cywilizacji reprezentowany przez myśl św. Tomasza z Akwinu.
Każdy kto choć pobieżnie zapoznał się z pracami autora tego eseju, publikowanymi na łamach Szturmu, wie że geny nie determinują ludzkiego zachowania, tylko na nie wpływają, gdyż każdy człowiek poza biologicznym dziedzictwem, został obdarzony wolą i odwagą, aby w heroicznym wysiłku stłumić w sobie to co złe a rozwinąć to co dobre.[29] Jak to już zostało zaznaczone powyżej, geny mogą sprawić że ludzie będą bardziej skłonni do alkoholizmu lub przemocy, ale żaden gen, lub kompleks genów nie może uczynić kogoś alkoholikiem - oznaczałoby to sprowadzenie ludzi do statusu genetycznych robotów, pozbawionych woli i odwagi do działania. Jednak stanowisko Jonesa w tej sprawie, całkowicie ignorujące opisywane w tym eseju wyniki badań naukowych, pozycjonuje katolicyzm na stanowisku kartezjańskiego „ducha w maszynie”
Najważniejszym elementem naszej odpowiedzi na ten rodzaj krytyki Etnicznego Nacjonalizmu, formułowanej przez katolickich intelektualistów(na których powołują się z kolei obywatelscy nacjonaliści) będzie całkowicie sofistyczny charakter zarzutów opartych rzekomo na scholastyce tomistycznej. Argument ten jest fałszywy, gdyż w nauczaniu katolickim zawsze była obecna idea, że dziedziczenie przyczynia się do moralnej konstrukcji człowieka. Jak pisze Karl Nemmensdorf, w nauczaniu katolickim człowiek jest doskonałym połączeniem dwóch odrębnych elementów: ciała i duszy. Ludzka dusza jest zarówno ożywczą zasadą ciała jak i nieśmiertelnym duchem.(dusze zwierząt nie są duchami i znikają wraz ze śmiercią). Dusze ludzkie mają zdolności duchowe, lub moce zwane intelektem i wolą. Intelekt ma siedzibę w duszy, ale jest zależny od zmysłów, zapewniających mu bodźców do działania. Do jakiego stopnia intelekt jest zależny od fizycznego mózgu, jest to problem, który od dawna fascynuje pokolenia katolickich teologów i psychologów. Odpowiedzi na pytanie o źródła zróżnicowania inteligencji u ludzi są dwie. Pierwsza polega na twierdzeniu o fizycznej jakości mózgu, jako źródle mocy i zasięgu działania intelektu. Druga opiera się na twierdzeniu że Bóg kształtuje duszę i jej intelekt zgodnie z budową ciała (i mózgu). Z obydwu tych wyjaśnień wynika wprost, że materialne ciało ma wpływ na duszę i jej możliwości[30]. Zresztą ostatnio K. Karoń, na początku swoich rozważań o „Historii antykultury” zwrócił uwagę na fałszywą dychotomię materia – duch (energia) [31], czego Kościół Katolicki zdawał się być świadomy przez całe wieki swojej historii, która to świadomość była szczególnie widoczna w dziełach Tomistów. Niemiecki przedstawiciel tego nurtu opat Vonier w swoim klasycznym dziele „Ludzka dusza” pisał : Wydaje się, że nie ma sprzeczności w przypuszczeniu, że dusze spirytualne mogą się znacznie różnić cechami, a Bóg formuje je zgodnie z różnicami dziedzicznych skłonności[.. . ]. Święty Tomasz z Akwinu wyraźnie skłania się ku poglądowi, że Wszechmogący Bóg kształtuje duszę, którą stwarza zgodnie z ciałem, w które ją wlewa. Tak długo, jak spirytualizm duszy jest chroniony, nie ma powodu, dla którego ciało ze swoimi cechami nie powinno być dla Boga okazją do stworzenia duszy o odpowiednich cechach.[ Biologiczna konstrukcja człowieka] ma swoje cechy i wady, których dusza nie może zmienić […] funkcją duszy jest[.. . ] dostroić wszystkie struny natury do najwyższej wysokości; ale całe strojenie na świecie nigdy nie zmieni marki instrumentu. [32] Te „rasistowskie”z punktu widzenia postsoborowego Kościoła pomysły prezentował również Jezuita Ernest R. Hull pisząc w „Formacji Charakteru”: Cielesne dary natury są [.. . ] nierównomiernie rozmieszczone; i stąd ogromna różnica jakości w składzie mózgu, nerwów, wrażliwych narządów i reszty. A ponieważ wszystkie nasze funkcje życiowe muszą być realizowane przez ten wspólny instrument zwany ciałem, wynika z tego, że ludzie powstają z ogromną początkową różnicą zdolności umysłowych, a nawet moralnych, zgodnie z cechami różnych organów. … Dlatego nie ma trudności z uznaniem faktu, że niektórzy ludzie rodzą się głupi, inni sprytni, niektórzy słabi, a inni silni, niektórzy ospali, inni żywiołowi i aktywni. Nawet w porządku moralnym są tacy, którzy niemal dosłownie rodzą się w ciele aniołów, podczas gdy inni są boleśnie podatni na gniew, lenistwo, obżarstwo[33]. Nawet w encyklopedii katolickiej z 1908 roku stanowisko wobec kwestii dziedziczenia miało podłoże ewolucyjne: Charakter jest wyrazem osobowości człowieka.. . . Charakter człowieka jest wypadkową dwóch odrębnych klas czynników: oryginalnych lub odziedziczonych elementów jego bytu i tych, które nabył. Z jednej strony, każda istota ludzka zaczyna się od określonej natury lub usposobienia - rodzimego wyposażenia zdolności do wiedzy, uczuć i tendencji do woli i działania - które różnią się w zależności od jednostki. To usposobienie zależy częściowo od struktury organizmu, a zwłaszcza od odziedziczonego przez niego układu nerwowego; po części być może także na stworzonej przez niego duszy.. . . Przeniesienie dziedzicznych dyspozycji z rodzica na potomstwo nie pociąga zatem za sobą konfliktu z doktryną stworzenia każdej ludzkiej duszy.
Dla człowieka nie ma nic ważniejszego niż jego charakter. Charakter jest najprawdziwszym przejawem tego, kim jest człowiek, bezpośrednim wyrazem jego moralnego temperamentu. I charakter pochodzi częściowo z dziedziczności. Opiera się na temperamencie i jest kształtowany przez zwykłe działanie woli, które decyduje między kierunkami działania. (Temperament może być uważany za naturalne skłonności osoby, przejawiające się zwłaszcza w jego osobowości, z silnym składnikiem dziedzicznym.) Charakter to „grupa wewnętrznych dyspozycji, wywodzących się z dziedziczności, środowiska, edukacji lub celowo ukształtowanych nawyków[34].
Na podstawie tych przykładów widać wyraźnie, że idea dziedziczenia genetycznego nie była związana z kościołem jedynie poprzez fakt, że jej odkrywca G. Mendel był katolickim zakonnikiem. Idea, że fizyczne, dziedziczne cechy ciała nadają człowiekowi niemałą część jego konstrukcji moralnej była istotnym elementem głównego nurtu tradycyjnej teologii katolickiej. Prawdziwa nauka katolicka nigdy nie gardziła materią, lecz ją wywyższała jako Boże dzieło twierdząc, że najwyższą formą materii we wszechświecie jest ciało ludzkie, ożywione przez duszę. Gdyby było inaczej Roman Dmowski nigdy zapewne, nie wyznając samemu tej pustynnej religii, nie „ochrzciłby”polskiego nacjonalizmu. W istocie, będąc ewolucjonistą, intuicyjnie pojmował religię jako kluczową adaptację do ekosystemu na poziomie wspólnoty etnicznej, co naukowo potwierdziły dopiero prace D. S. Wilsona i J. Heidta [35] sto lat później.
Natomiast pozorni radykałowie katoliccy, którzy podobnie jak obywatelscy nacjonaliści dostrzegając rolę mniejszości etnicznych oraz postmodernizmu w niszczeniu religii, pisząc jednocześnie, że „bez kościoła katolickiego Europa przypominałaby obecnie Somalię”[36] reprezentują w istocie jedynie wcześniejsze stadium tej samej postsoborowej choroby, która zniszczyła myśl katolicką głównego nurtu.
Kończąc powyższą pracę, należy podsumować wyłaniające się z niej wnioski, aby dzięki temu móc wyznaczyć kierunki adaptacyjnego działania dla każdego nacjonalisty.
Orędownicy obywatelskiego nacjonalizmu nie są naszymi towarzyszami walki. Będąc genetycznymi mutantami, poprzez swoje życiowe wybory, skazali również swoje dusze na zatracenie, co z pewnością w eliminuje ich dziedzictwo z naszego etnosu w każdym egzystencjalnym aspekcie.
Natomiast my etniczni nacjonaliści musimy podążać w jedności, realizując prawo natury na każdym poziomie naturalnej selekcji i doboru płciowego. Zadania, które kiedyś nasi przodkowie realizowali intuicyjnie, dzisiaj musimy uczynić elementem zbiorowej świadomości. Wobec faktu, że nasi wrogowie wykorzystują intuicję, znaczniki kulturowe oraz instytucje dystrybucji autorytetu w celu zniszczenia naszych genetycznych interesów, nie mamy innego wyjścia. Na szczęście prawda, którą cały czas odkrywa nauka jest dla nas jako oręż coraz bardziej dostępna. Dzięki niej stwarzajmy własne portfele dostosowania aby na każdym etapie życia pomnażać i zabezpieczać nasze etniczne interesy genetyczne. Niech instytucje dystrybucji autorytetu, które są w naszych rękach, promują odpowiednie wzory postępowania adekwatne do wieku płci i innych indywidualnych cech. Bowiem wbrew liberalnym bredniom nasza indywidualna egzystencja bez społecznego kontekstu sama w sobie nie ma żadnej istotnej wartości. Parafrazując Dmowskiego dopiero to, że identyfikujemy się jako Polacy nadaje naszemu życiu podstawowy sens, polegający na realizacji obowiązków wynikających z praw natury i ewolucji. Im wyższa jest nasza świadomość, wola tworzycielska i biologiczne możliwości organizmu tym więcej sposobów na zwiększenie dostosowania swojej grupy etnicznej znajdzie każdy z nas. Tak więc dopiero fakt, w jak wielu społecznych, ekologicznych, ekonomicznych i innych kontekstach na przestrzeni życia jesteśmy w stanie zwiększać interes genetyczny naszego etnosu i podgatunku dowodzi jak wysoki typ człowieka i nacjonalisty przedstawiamy.
Zgodnie z tym ewolucyjnym algorytmem każdy z nas, będąc młodym człowiekiem powinien zwiększać swoje fitness zarówno wspierając rodzeństwo i rodziców, jak i dbając o własny rozwój z jednej strony, a z drugiej angażując się w możliwie najbardziej elitarne organizacje nacjonalistyczne aby tam realizować altruizm wobec etnosu jednocześnie monitorując towarzyszów i towarzyszki pod kątem ich eugenicznej jakości. Działalność ta jak i wzajemne monitorowanie ma bowiem kluczową funkcje dla kolejnego etapu życia każdego nacjonalisty którym jest dobór płciowy i reprodukcja na możliwie wysokim poziomie, tak aby z jednej strony zniwelować ujemny przyrost naturalny całego etnosu a z drugiej eugenicznie go ulepszyć, powodując,ż e tam gdzie znajdują się allele kodujące zarówno pozytywny jak i negatywny etnocentryzm będzie on wyższy niż średnia.
Wreszcie w sile wieku każdy z nas może odegrać największą rolę w największej liczbie kontekstów. Mając ugruntowaną pozycję materialną można z większą skutecznością i na większą skalę realizować dobroczynny altruizm wobec innych aktywistów mających materialne problemy, służyć radą, pomocą i doświadczeniem, zwiększając dostosowanie młodzieży a co najważniejsze mając kilkunastoletnie doświadczenie i rozeznanie w środowisku nacjonalistycznym, skłaniać swoje potomstwo do małżeństwa z potomstwem innych aktywistów tak aby geny etnocentryzmu stawały się dominujące w genomie kolejnych generacji.
Cały czas warto jednakowoż utrzymywać sprawność fizyczną aby w kluczowym momencie życia kiedy każdy z nas w obliczu rychłej śmierci spowodowanej np. nieuleczalną chorobą mógł poświęcić swoje życie w akcie ofiarnego altruizmu powstrzymując tym samym tych którzy w drodze imigracji chcą zniszczyć genetyczny interes naszego etnosu i podgatunku.
Załącznik
Współczynnik pokrewieństwa i Oddzielone Populacje
Streszczenie
Współczynnik pokrewieństwa między dwoma organizmami diploidalnymi opisuje ich ogólne podobieństwo genetyczne względem siebie w stosunku do pewnej populacji podstawowej. Na przykład pokrewieństwo między rodzicem a potomstwem 1/4 opisuje dzielenie genów w większym stopniu niż losowe dzielenie w losowej populacji krycia. W podzielonej populacji statystyka Fst opisuje dzielenie genów w podgrupach w ten sam sposób. Ponieważ Fst wśród populacji ludzkiej w skali światowej wynosi niezawodnie 10 do 15 procent, pokrewieństwo między dwiema osobami tej samej populacji ludzkiej jest równoznaczne z pokrewieństwa między dziadkiem i wnukiem lub między przyrodnim rodzeństwem. Powszechne twierdzenie, że jest to wartość niewielka i nieistotna, powinno zostać ponownie zbadane.
Współczynnik pokrewieństwa
Łatwo jest zrozumieć, dlaczego opieka rodzicielska rozwinęła się w wielu liniach: rodzice i potomstwo dzielą geny, tak że wysiłek rodzicielski poświęcony potomstwu jest w rzeczywistości wysiłkiem poświęconym genom rodziców. Hamilton (1964) sformalizował ten pogląd i rozszerzył go na dowolne stopnie pokrewieństwa. Kiedy Hamilton i inni opisywali teorię, często wyrażali się w kategoriach tożsamości genów poprzez pochodzenie, myśląc na przykład o połowie genów nuklearnych w diploidalnym potomstwie, które są identyczne z genami u rodzica. Wielu autorów mówiło także o wspólnych genach. Żaden z tych opisów nie jest całkowicie dokładny. Mogę na przykład dzielić wiele genów z cebulą, ale ten podział genów nie ma znaczenia dla ewolucji zachowań społecznych u ludzi.
Lepszym sposobem na myślenie o pokrewieństwie, związku i teorii Hamiltona jest myślenie o dzieleniu genów w nadmiarze i deficycie przypadkowego dzielenia genów. Rodzic dzieli o wiele więcej niż połowę swoich genów z potomstwem, ale w losowej populacji krycia połowa tych genów jest z pewnością identyczna, ponieważ pochodzą od rodzica, podczas gdy dzielenie genów z drugą połową genomu dziecka jest tym, co jest wspólne z każdym losowym członkiem populacji.
Podczas gdy Hamilton pisał swoją teorię w kategoriach stopnia pokrewieństwa, większość genetyków populacyjnych rozumuje zamiast tego w kategoriach współczynnika pokrewieństwa. Po poznaniu pokrewieństwa związek natychmiast powstaje z prostej formuły (Bulmer 1994).
Oto definicja pokrewieństwa między osobą x a osobą y: wybieramy losowy gen w locus od x i niech częstotliwość w populacji tego genu będzie p. Teraz wybieramy gen z tego samego locus od osoby y. Prawdopodobieństwo tego, że gen w y jest taki sam jak gen wybrany z x, czyli Py jest
Py = Fxy + (1 – Fxy)P
Interpretacja tego jest taka, że z prawdopodobieństwem F geny są takie same, z prawdopodobieństwem 1 - F są różne, w którym to przypadku prawdopodobieństwo tożsamości to tylko częstotliwość populacji p (Harpending 1979). Zmiana równania daje definicję współczynnika pokrewieństwa:
Fxy = (Py -P)/(1 – P) (1)
Współczynniki pokrewieństwa w losowo dobranej populacji diploidalnej są proste i dobrze znane. Na przykład wybierzmy gen ode mnie, a następnie wybierzmy ode mnie inny gen z tego samego locus. Z prawdopodobieństwem 1/2 wybraliśmy ten sam gen, natomiast z prawdopodobieństwem 1/2 wybraliśmy inny gen w tym locus. Dlatego prawdopodobieństwo, że drugi gen jest taki sam jak pierwszy, wynosi tylko 1/2 + p / 2, a podstawienie tej częstotliwości warunkowej we wzorze na pokrewieństwo pokazuje, że moje pokrewieństwo z samym sobą wynosi tylko 1/2. To samo rozumowanie prowadzi do dobrze znanych wartości 1/4 dla mojego dziecka, 1/8 dla mojego wnuka, mojego przyrodniego rodzeństwa lub mojego siostrzeńca i tak dalej.
Bardzo ważne jest, aby współczynnik pokrewieństwa nie był mylony ze stopniem pokrewieństwa [ Hamiltonowskim ]. Są to różne koncepcyjnie i liczbowo twory. Stopień pokrewieństwa można uznać za „część wspólnych genów” między dwoma organizmami. Pokrewieństwo to jest znane wielu biologom, odkąd W. D. Hamilton opracował swoją słynną teorię selekcji krewniaczej pod względem stopnia pokrewieństwa. Jednak większość późniejszych opracowań teorii dotyczyła współczynników pokrewieństwa.
W losowej, diploidalnej populacji krycia, związek między dwoma współczynnikami jest prosty: stopień pokrewieństwa jest tylko dwa razy większy niż współczynnik pokrewieństwa. Ta prosta reguła kciuka załamuje się, gdy tylko pojawią się jakiekolwiek komplikacje, takie jak chów wsobny lub struktura populacji. Najlepsza ogólna definicja współczynnika pokrewieństwa Rxy między osobami x i y to (Bulmer 1994):
Rxy = Fxy / Fxx
gdzie Fxy oznacza pokrewieństwo między x I y, a Fxx oznacza pokrewieństwo x z samym sobą. Ma to tę interesującą właściwość, że niekoniecznie są one symetryczne: Rxy ogólnie nie jest równe Ryx.
Oddzielone Populacje
Większość zastosowań teorii Hamiltona w biologii wykorzystuje pokrewieństwo i relacje wynikające z relacji genealogicznych. Na przykład, uważamy, że opieka rodzicielska ewoluuje, ponieważ rodzice i potomstwo dzielą geny. Ale dzielenie genów (zawsze więcej niż losowe dzielenie genów) może wystąpić w innych sytuacjach. W podzielonej populacji jednostki dzielą geny z innymi członkami tego samego demu, a te wspólne geny są paliwem dla ewolucji dzięki działaniu dostosowania łącznego w dokładnie taki sam sposób, w jaki relacje rodowodowe, takie jak między rodzicem a dzieckiem, są paliwem dla ewolucji poprzez działanie dostosowania łącznego.
Wyliczamy w tym miejscu współczynnik pokrewieństwa między oddzieloną populacją a stopniem pokrewieństwa w bardzo prostym przypadku, ale formuły te mają zastosowanie znacznie bardziej ogólnie, niż sugeruje to nasze proste wyprowadzenie. W tym miejscu muszę wspomnieć, że nasze pochodne dotyczą dużych populacji. W przypadku małych grup („grup cechowych”, jak je nazywa D. S. Wilson) musielibyśmy wziąć pod uwagę, że jeśli wybieramy gen od osobnika, częstotliwość tego genu w pozostałej części puli genów demu jest nieznacznie zmniejszona. Dokładne traktowanie małych demów prowadzi do nieznośnych algebraicznych warunków rzędu 1 / n, gdzie n jest rozmiarem dema. Jednak rozważmy duże grupy ignorując te warunki.
Rozważmy populację złożoną z dwóch demów dokładnie tej samej wielkości i locus genetycznego z dokładnie dwoma allelami. Wniosek z algebry poniżej jest taki, że znana statystyka opisująca podział populacji, Fst jest właśnie pokrewieństwem między członkami tego samego demu. Innymi słowy różnice genetyczne między demami wskazują na podobieństwo genetyczne w obrębie demów, a Fst jest tylko współczynnikiem pokrewieństwa między członkami tego samego demu ze względu na strukturę populacji. Na przykład Fst wśród populacji ludzkich wynosi około 1/8, a jest to tylko współczynnik pokrewieństwa w jednej populacji między dziadkiem i wnukiem, wujem i siostrzeńcem lub dwojgiem przyrodniego rodzeństwa. W zróżnicowanym świecie członkowie tej samej populacji są ze sobą spokrewnieni w takim samym stopniu, w jakim dziadkowie i wnuki są spokrewnieni w jednej populacji.
Istnieją dwa geny o równej wielkości oznaczone A i B. W locus częstotliwość genu to pA w demie A i pB w demie B. Częstotliwościami w dwóch demach alternatywnego allelu są qA i qB. Ogólne średnie częstotliwości to po prostu p i q. Do opisania częstotliwości genów wygodnie jest użyć nieco innej notacji:
pA = p +σ
pB = p – σ
więc oczywiście:
qA = q – σ
qB = q + σ
Teraz wyobraźmy sobie, że wybieramy losowo gen z populacji, a następnie wybieramy inny gen z tego samego locus z tego samego demu. Jaki jest współczynnik pokrewieństwa między demami? Aby to znaleźć, używamy powyższego wzoru (1).
Z prawdopodobieństwem 1/2 początkowo wybieramy kogoś z populacji A, a z prawdopodobieństwem pA wybieramy allel, którego częstotliwość wynosi pA · Z prawdopodobieństwem qA = 1 - pA wybieramy alternatywny allel. Umieszczając te możliwości w równaniu (1) mamy
F = (1/2)pA(pA – p)/q + (1/2)pB(pB-p)/q + (1/2)qA(qA-q)/p + (1/2)qB(qB-q)/p
Przy użyciu wymienionych wyżej podstawień, otrzymujemy:
F = {(p+б)(б) + (p-б)(-б)}/2q + {(q-б)(-б) + (q+б)(б)}/2p
= 2б²/2q + 2δ²/2p
I momentu gdy p + q = 1
F=4δ²/4pq
= δ²/pq
Jest to po prostu odległość genetyczna Fst między dwiema populacjami - wariancja częstotliwości genu podzielona przez średnią częstotliwość genu pomnożoną przez jej dopełniacz. Gdy Fst jest raportowane dla zbioru populacji, jest to w rzeczywistości średnia wszystkich statystyk Fst dla całej populacji. Statystyka jest obliczana dla każdego allelu w każdym locus, a następnie uśredniana dla wszystkich loci.
Wiele badań potwierdza, że Fst w próbkach populacji ludzkiej na świecie wynosi od dziesięciu do piętnastu procent. Jeśli uwzględniono małe, długo izolowane populacje, liczba ta jest zwykle nieco wyższa. Konserwatywna ogólna liczba dla naszego gatunku to Fst ≈ 0,125 = 1/8. Liczbę tę podał Cavalli-Sforza w 1966 r., A szeroko cytowany artykuł Lewontina (1972) długo argumentował, że jest to niewielka liczba, sugerując, że różnice w populacji ludzkiej są trywialne. Alternatywną perspektywą jest to, że pokrewieństwo między dziadkami i wnukami, równoważne pokrewieństwu w ludzkich populacjach, nie jest tak trywialne. W celu dalszej dyskusji patrz Klein i Takahata (2002, s. 3 87–390).
Pokrewieństwo w Oddzielonej Populacji
Równanie (1) i jego wyprowadzenie pokazuje, że jeśli wybieramy gen losowo z populacji dwóch demów, odkryjemy, że jego ogólna częstotliwość wynosi p, wówczas częstotliwość tego genu w tym samym demie [z którego był gen] jest średnio
psam = p + (1 – p)Fst
podczas gdy częstotliwość tego genu w drugim demie wynosi średnio:
P drugiego= p - (1 – p)Fst.
Stosując równanie (1) i te relacje, możemy łatwo wyliczyć współczynniki pokrewieństwa i stopień pokrewieństwa w obrębie demu, oraz pomiędzy demami.
Współczynnik pokrewieństwa jednostki z kimś z jego własnego demu wynosi tylko Fst, podczas gdy jego pokrewieństwo z kimś z innego demu wynosi - Fst. A co z pokrewieństwem ze samym sobą w podzielonej populacji? Wybierzmy gen od osoby, a następnie losowo wybierzmy inny gen od tej samej osoby: z prawdopodobieństwem 1/2 wybraliśmy ten sam gen i z prawdopodobieństwem 1/2 wybraliśmy inny, w którym to przypadku prawdopodobieństwo, że to jest ten sam, wynosi p + (1 - p) Fst
Dlatego:
P samego siebie = (½)(1 + p + (1 – p)Fst)
Używając równania (1) otrzymujemy :
F samego siebie = (1 + Fst)/2
zamiast zwykłego 1/2 pokrewieństwa ze sobą w jednej losowej populacji krycia.
Łatwo jest uzyskać znane współczynniki pokrewieństwa rodzinnego w ten sam sposób: na przykład pokrewieństwo z dzieckiem, gdy drugi rodzic pochodzi z tego samego demu:
F dziecka = 1 /4 + 3 Fst/4
i tak dalej. Zasadniczo, jeśli pokrewieństwo w losowej populacji krycia z krewnym wynosi 1 / x, wówczas w podzielonej populacji pokrewieństwo z tym samym krewnym wynosi:
F krewnego stopnia x = 1 /x + (1 - x)Fst /x (2)
A co w przypadku współczynnika pokrewieństwa z krewnym, który jest hybrydą między populacyjną? Rozważmy na przykład dziecko, którego drugi rodzic pochodzi z drugiego demu. Wybierzmy gen od rodzica: prawdopodobieństwo wybrania tego samego genu od dziecka wynosi 1/4, prawdopodobieństwo wybrania genu od dziecka nie jest identyczne z pierwszym, ale z tego samego demu, co rodzic wynosi 1/4, i prawdopodobieństwo wybrania genu z drugiego demu wynosi 1/2. Podsumowując, prawdopodobieństwo znalezienia tego samego genu wynosi :
P potomstwa hybrydowego = 1/4 + 1/4(p + (1 - p)Fst) + 1/2(p - (1 – p)Fst).
Używając równania (1) otrzymujemy :
F potomstwa hybrydowego = 1/4 – Fst/4.
Zasadniczo te same pochodne pokazują, że pokrewieństwo z hybrydowym krewnym stopnia x, co oznacza krewnego, z którym pokrewieństwo w losowej populacji krycia wynosi x, jest
F hybrydowego krewnego stopnia x = 1/x – Fst/x (3)
Różnica między równaniami (2) i (3) to po prostu Fst, czyli różnica między współczynnikiem pokrewieństwa z krewnym wewnątrz demowym a krewnym hybrydowym. Zauważmy również, że gdy x staje się duże, równanie (2) pokazuje, że współczynnik pokrewieństwa z losowym członkiem tego samego demu to Fst, a współczynnik pokrewieństwa z innym,niepowiązanym stopniem pokrewieństwa potomstwem hybrydowym wynosi 0.
prof. Henry Harpending
Literatura:
Bulmer, M. (1994).”Theoretical evolutionary ecology”. Sinauer, Sunderland, Massachusetts.
Cavalli-Sforza, L. L. (1966). „Population structure and human evolution”. Proceedings of the Royal Society of London B, 164, 362-79.
Hamilton, W. D. (1964). „The genetic evolution of social behavior”, części 1 i 2
Journal of Theoretical Biology, 7, 1 -51. Dodatek 1333
Harpending, H. (1979). „The population genetics of interactions”. American Naturalist,113, 622-30.
Klein, J. Takahata, N. (2002). „Where do we come from ? The molecular evidence
for human descent”. Springer, Berlin.
Lewontin, R. C. (1972). „The apportionment of human diversity.” Evolutionary
Biology, 6, 3 8 1 -98.
[1] . Paradoksalnie, rozgałęziające się pokrewieństwo w formie drzewa genealogicznego jest trafnym intuicyjnie zobrazowaniem rzeczywistości genetycznego pokrewieństwa pomiędzy populacjami. W przypadku rodowodów pojedynczych ludzi wewnątrz homogenicznej populacji rzeczywistość jest sprzeczna z konceptualizacją pokrewieństwa jako wciąż rozgałęziającego się drzewa gdyż w tym przypadku mamy ciągły splot, ową mitologiczną tkaninę pokrewieństwa w którym rozgałęziająca się linia, po kilku stopniach pokoleniowych splątuje się ze sobą ponownie utrzymując stężenie pokrewieństwa na średnim stałym poziomie w całej populacji
[2] .Te fascynujące odkrycia etnografów, ukazujące wiarę Słowian w częściową reinkarnację ludzi we własnych wnukach, przy jednoczesnym odejściu w zaświaty drugiej części duszy związanej w wolną wolą, ukazują scholastyczne korzenie indoeuropejskiej tradycji naukowej, która w przeciwieństwie do religii pustynnych, starała się rozumowo badać religię na drodze badania argumentów empirycznych. Dzięki temu nasza cywilizacja stworzyła na bazie religii naukę, której poszczególne gałęzie (w tym przypadku genetyka behawioralna)potwierdzają, intuicyjne wyobrażenia religijne naszych przodków.
[3] Samo badanie rodowodów ludzkich jest pomocne jedynie w przypadku dziedziczenia chromosomu y (tzw. haplogrupy) w przypadku mężczyzn oraz dna mitochondrialnego (pozanuklearnego) dna w przypadku kobiet co będąc pomocnym dla badania rodowodów, staje się problematyczne dla genetyki behawioralnej (np X-DNA - umożliwia badanie pokrewieństwa tylko w niektórych liniach, ponieważ dziedziczony jest inaczej niż inne chromosomy (mężczyźni dziedziczą jeden z chromosomów X swojej matki, a kobiety po jednym od każdego rodziców; przy czym nie zawsze zachodzi rekombinacji) i może się zdarzyć, że osoby o bliskim stopniu pokrewieństwa (brat i siostra lub dwaj bracia nie będący bliźniakami jednojajowymi) odziedziczą po matce różne chromosomy X
[4] . D.L.Hartl, A.G.Clark „Podstawy genetyki populacyjnej”Warszawa 2009 s.43
[5] . Tamże s. 44
[6] . F. Salter „On genetic interests. Family. Ethnicity. And Humanity in an Age af Mass Migration”New Brunswick 2007 s. 46
[7] . J.P. Ruschton „Race. Evolution and behaviour”New Brunswick 1997 s. 81
[8] . F. Salter „On genetic...”s.47
[9] . Całą debatę na ten temat, pomiędzy C. Coonem a A. Montagu (właść. Israel Ehrenberg) i jej znaczenie opisuje V.Sarich. Zob. V.Sarich, F. Mielle „Race. A reality of human differences”s.105-112
[10] .Dryf genetyczny powoduje, że pokrewieństwo między losowymi członkami grupy zwiększa się w każdym pokoleniu w tempie 1/2N, gdzie N jest populacją lęgową (Cavalli-Sforza i Bodmer 1971/1999, s. 707). Załóżmy, że rodowe grupy ludzkie były tak duże, jak współczesne im plemiona i grupy łowców-zbieraczy, około 5OOosób, każda z 300 reprodukcyjnymi osobnikami, tak że N = 300. Wtedy pokrewieństwo wewnątrz pokolenia wzrosło o około 0,0017. Załóżmy, że czas generacji wynosi 25 lat. Następnie co tysiąc lat pokrewieństwo wzrastało o około 0,067, czyli nieco więcej niż pokrewieństwo pierwszych kuzynów. Przy takim tempie, w ciągu zaledwie 3,75O lat, odległość genetyczna między oddzielonymi populacjami byłaby tak duża, że losowi członkowie grupy mieliby pokrewieństwo 0,25 czyli odpowiadające pokrewieństwu między między rodzicem a dzieckiem.
[11] .www.theoccidentalobserver.net%2F2019%2F11%2F24%2Fcan-church-influence-explain-western-individualism-comment-on-the-church-intensive-kinship-and-global-psychological-variation-by-jonathan-f-schulz-et-al%2F&anno=2&prev=search
[12] .Jak pisze Heidt na przykład klimat Afryki był bardzo zmienny w okresie od 140 000 do 70 000 lat temu. Za każdym razem, gdy klimat zmieniał się z cieplejszego na bardziej chłodny albo z wilgotnego na suchy, ludzie musieli szukać nowych źródeł pożywienia i – jak można przypuszczać – często cierpieli zabójczy głód. Katastrofalny w skutkach wybuch wulkanu Toba w Indonezji, do którego doszło 74 000 lat temu, mógł radykalnie zmienić klimat na Ziemi w ciągu zaledwie roku. Niezależnie od przyczyny wiemy, że w którymś momencie prawie wszyscy ludzie wyginęli, a jednocześnie mogło to być impulsem dla pewnych grupek do pierwszych migracji poza afrykańską sawannę.
Niezależnie od tego, czy erupcja ta zmieniła bieg ewolucji naszego gatunku, należy zaznaczyć że ewolucja nie jest procesem płynnym i stopniowym, jak się zakłada w większości symulacji komputerowych. Prawdopodobnie obfituje ona w zdarzenia krytyczne (black swan events) – nieprzewidywalne, mało prawdopodobne zdarzenia opisane przez N. Taleba, które udaremniają nasze próby modelowania procesów przy użyciu zaledwie kilku zmiennych i paru założeń opartych na „normalnych” warunkach. Zob. Taleb, N.. Czarny łabędź. O skutkach nieprzewidywalnych
zdarzeń. Warszawa: (2014)
[13] .Plemiona, w których ludzie spędzili tak wiele ze swojej historii, były oparte o granice terytorialnie i pilnowały ich. To samo dotyczy współczesnych państw. Szczególną cechą bronionego terytorium jest to, że izoluje ono ludność od perypetii zaburzeń demograficznych w metapopulacji, a mianowicie od związanych z nią zjawisk nierównomiernego wzrostu populacji i migracji. Kiedy etnos kontroluje granice terytorium, które jest wystarczająco duże, aby utrzymać populację, utrata liczb w stosunku do innych etnosów niekoniecznie jest śmiertelna, tj. Nie musi prowadzić do wymiany ludności. Odpowiednio bronione terytorium gwarantuje ciągłość i szansę na przetrwanie chwilowego spadku liczebności. F. Salter „On genetic...”s. 60
[14] .https://pl.wikipedia.org/wiki/Tragedia_wsp%C3%B3lnego_pastwiska
[15] . E.O. Wilson „Pół Ziemi. Walka naszej planety o życie”Warszawa 2017 s.68
[16] . Imigranci muszą wpłynąć na zdolność kraju do utrzymania ludności tubylczej. Jeśli imigranci wnoszą wkład w gospodarkę w sposób, którego ludność tubylcza nie jest w stanie wnieść, zwiększa się nośność. Jeśli wyczerpują zasoby lub średnią produktywność, obniżają tę pojemność, zastępując miejsce potencjalnie urodzonych tubylców. Salter „On genetic..”s.62
[17] .Tamże s.66
[18] Realny wzorzec zastępowalności pokoleń występujący w niektórych społeczeństwach zachodnich, ze względu na połączenie liczby urodzeń poniżej zastępowalności i imigracji z odległych genetycznie populacji jest przez połączenie tych danych zakłamany. Typowy wzorzec spadku liczby urodzonych tubylców z powodu nadmiaru zgonów nad urodzeniami jest w istocie tragiczny, maskuje go względna stabilność ogólnej populacji spowodowana imigracją. Dotyczy to następujących krajów (z przekroczeniami liczby zgonów w porównaniu do liczby urodzeń rdzennej populacji w nawiasach): Włochy (11 procent), Szwecja (7 procent), Niemcy (14 procent) i Austria (0,6 procent) Zob. CIA World Factbook, January 2002, http://www.cia.gov/cia/publications/factbook/.
[19] .http://szturm.com.pl/index.php/miesiecznik/item/1044-jaroslaw-ostrogniew-pandemia-koronawirusa-ostateczny-upadek-globalizmu
[20] .W społeczeństwach wieloetnicznych niski poziom zaufania istnieje nie tylko pomiędzy mniejszościowymi grupami etnicznymi, a grupą większościową, ale również w samej grupie większościowej, która żyje na swoim terytorium z mniejszościami. Potwierdzają to także badanie Alesina i La Ferrera(2002). Jak zaznacza E.Dutton, jedną z możliwości jest to że istnieje teraz ryzyko że każdy członek większościowej grupy etnicznej może zdradzić i zwiększyć swój status poprzez koalicję z mniejszościami. Innym wytłumaczeniem tego zjawiska jest to, że członkowie grupy większościowej, widząc jak ich rodacy zezwolili na wtargnięcie na ich terytorium, wnioskują że dłużej nie można im ufać E. Dutton „Race diferences in etnocentrism”London 2016 s.233
[21] . Tamże s. 235 Co ciekawe do tych samych wniosków doszedł amerykański badacz R. Putnam przeanalizował zasoby kapitału społecznego w setkach amerykańskich społeczności i doszedł do wniosku, że wysoki poziom imigracji i różnorodności etnicznej wydaje się powodować spadek kapitału społecznego. Był on jednak bardziej wnikliwy (jak cała jego 500 stronicowa „samotna gra w kręgle „będąca porażającym studium społecznego upadku USA) jego kwestionariusz umożliwiał rozróżnienie dwóch rodzajów kapitału społecznego: kapitał pomostowy (bridging capital) to zaufanie między grupami, pomiędzy ludźmi o różnych wartościach i tożsamościach, podczas gdy kapitał wiążący (bonding capital) to zaufanie w obrębie danej grupy. Putnam odkrył, że różnorodność pomniejsza oba rodzaje kapitału. Oto jego wniosek:Różnorodność wydaje się wyzwalać nie podział na grupę własną i obcą, lecz anomię lub izolację społeczną. Mówiąc potocznie, ludzie żyjący w środowisku zróżnicowanym etnicznie wydają się kulić w sobie – chowają się w skorupie jak żółwie.
[22] .Na podstawie tego wskaźnika (heterogeniczność etniczna- EH) Vanhannen zbadał różnice w poziomie konfliktu etnicznego (EC)w oparciu o skalę 1-5. (1) o niskiej wartości i (5) o wysokiej wartości. Kategorie Vanhanen to: (1) Drobne incydenty na poziomie lokalnym, niewielkie etniczne partie polityczne lub organizacje interesów. (2) Znacząca lokalna przemoc na tle etnicznym, znaczące partie lub grupy interesów oparte na pochodzeniu etnicznym, zinstytucjonalizowana dyskryminacja etniczna. (3) Gwałtowny konflikt etniczny, separatystyczne dążenia w niektórych częściach kraju, ważne etniczne partie polityczne i grupy interesów, poważna dyskryminacja ujarzmionej grupy etnicznej. (4) Wojna domowa, bunt etniczny, terroryzm, wojny separatystyczne, etniczne partie polityczne / grupy interesów dominują w polityce, tłumione są duże grupy etniczne, uchodźcy etniczni. (5) Gwałtowna wojna domowa na tle etnicznym dominująca w polityce, ludobójstwio na tle etnicznym.
[23] .Na podstawie 176 krajów(KE) koreluje z demokratyzacją na poziomie -0,22, ze Wskaźnikiem Rozwoju Społecznego 2010 na poziomie -0,39 oraz z PKB / PNB na mieszkańca na poziomie -0,253. Sensowne jest, że konflikt etniczny byłby związany z obniżonym standardem życia z wielu powodów. Wysoki poziom konfliktu etnicznego spowodowałby, że społeczeństwo byłoby niebezpieczne i niestabilne, a tym samym zniechęciłoby się co do zaangażowania w projekty długoterminowe, co znalazłoby odzwierciedlenie w braku współpracy i zaufania w społeczeństwie, które zasadniczo uwikłane byłoby w wojnę o różnym nasileniu. Z tego powodu można się spodziewać, że życie w wysoce zróżnicowanym etnicznie społeczeństwie byłoby niezwykle stresujące, a ponieważ etnocentryzm wydaje się wysoce instynktowny, takie stresy podniosą zarówno międzygrupowy (wewnątrz kraju) etnocentryzm i etnocentryzm grupowy, co czyni takie społeczeństwa stosunkowo wrogimi wobec obcokrajowców. E. Dutton „Race..”s. 238
[24] .Papież Franciszek domaga się życzliwego i otwartego przyjmowania muzułmańskich imigrantów oraz stwierdza że „w obliczu niosących niepokój okresów gwałtownego fundamentalizmu nasz szacunek wobec prawdziwych wyznawców islamu powinien doprowadzić nas do tego by unikać złowieszczych uogólnień, gdyż autentyczny islam i właściwe odczytanie Koranu sprzeciwiają się wszelkiej formie przemocy”. Doprawdy nie wiadomo jak rozumieć te słowa. Jedno jest pewne, że nie mają one nic wspólnego z faktami i albo świadczą o dramatycznej utracie kontaktu z rzeczywistością albo są wyrazem jakiegoś nieprawdopodobnego wręcz utopizmu.P. Lisicki „Dżihad i samozagłada Zacsodu”Lublin 2015 s.96
[25] . T. Gabiś „Z zapisków lat 1947 – 51”Stańczyk nr 2(19) 1993 s.34
[26] . Carl Schmitt „Teologia polityczna i inne pisma”W-wa 2012 s.258-259. W duchu rozważanego w tym eseju etnicznego interesu genetycznego, możemy sparafrazować przywołanego przez Schmitta E.Forcelliniego pisząc, że wróg to ten który zagraża genetycznym interesom naszego etnosu, czym różni się od nieprzyjaciela z którym mamy osobistą waśń wynikającą z rywalizacji o zasoby lub różnicy zdań. Można ich tak odróżnić, bo nieprzyjaciel nas nienawidzi chcąc zmniejszyć nasze dostosowanie łączne, a wróg chce zniszczyć nasz genetyczny interes atakując nasz etnos i państwowe granice.
[27] .Michael E. Jones, autor „Żydowskiego ducha Rewolucji” zyskał rozgłos dzięki swojej publicystyce która charakteryzuje się zawierania złożonych procesów społeczno – kulturowych w jednym zdaniu np. „Nowoczesność to zracjonalizowane niewłaściwe zachowanie seksualne”. Życiorys tego pisarza, którego kariera zaczęła się w momencie zwolnienia go z katolickiego St. Marys College w Soutch Bend, za jednoznaczny sprzeciw wobec aborcji, jest najlepszym świadectwem kondycji moralnej współczesnego establishmentu kościelnego.
[28] https://www.theoccidentalobserver.net/2019/12/10/ethnos-needs-logos-and-genos/
[29] . Securius nr 1/ 1996
[30] .https://www.theoccidentalobserver.net/2019/12/10/ethnos-needs-logos-and-genos/
[31] . K. Karoń „Historia antykultury”Warszawa 2018 s. 20
[32] . Vonier, Abbot. The Human Soul. Maryland, 2010. s.45 - 47
[33] . Hull, Ernest R., SJ The Formation of Character. cyt za https://www.theoccidentalobserver.net/2019/12/10/ethnos-needs-logos-and-genos/
[34] . Attwater, Donald. A Catholic Dictionary. New York 1943 s.96 https://www.newadvent.org/cathen/03586a.htm
[35] . Zob zwłaszcza D. S. Wilson „Darwins Catedral. Evolution, Religion and the nature of society”Chicago 2002
[36] . Średnia inteligencja rdzennych Europejczyków wynosi 100IQ natomiast IQ rdzennych Somalijczyków to 68 R. Lynn „Race differrences in inteligence. An evolutionary analysis”Augusta 2006 s.22 – 26 Złowrogim dowodem na fałszywość tezy Jonesa że to kultura (religia) tworzy etnos jest Synod Amazoński i jego deklaracje, ukazujące że po wiekach od zaszczepienia indoeuropejskiej tradycji zawartej w katolicyzmie na drastycznie odmienny rasowo grunt południowoamerykański, w wyniku koewolucji kulturowo genetycznej, to nie katolicyzm zmienia rasę Indian, tylko rasa zmienia katolicką tradycję z indoeuropejskiego realizmu naukowego na tropikalny szamanizm